Bourhis Jean-Marie, Receveur-Bréchot Véronique, Oglesbee Michael, Zhang Xinsheng, Buccellato Matthew, Darbon Hervé, Canard Bruno, Finet Stéphanie, Longhi Sonia
Architecture et Fonction des Macromolécules Biologiques (AFMB), UMR 6098 CNRS at Universités Aix-Marseille I et II, ESIL, Campus de Luminy, 13288 Marseille Cedex 09, France.
Protein Sci. 2005 Aug;14(8):1975-92. doi: 10.1110/ps.051411805.
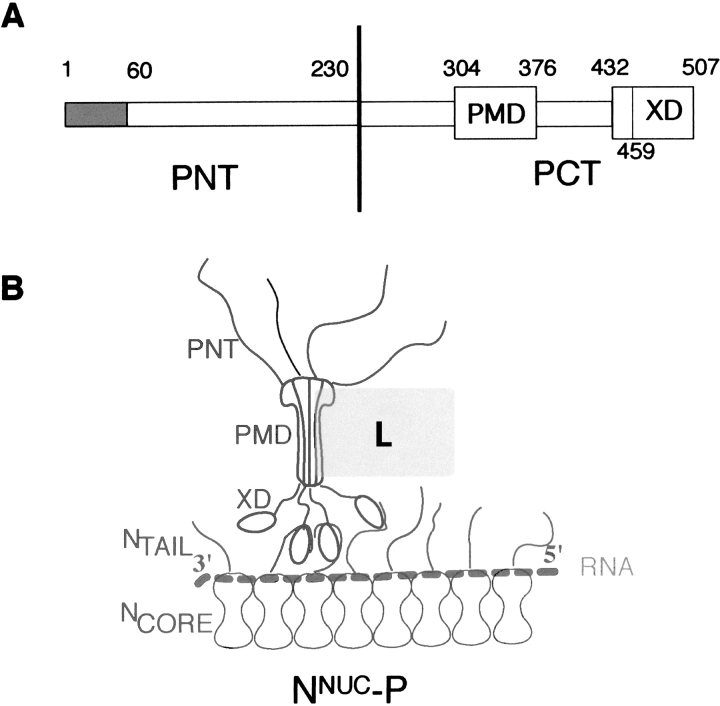
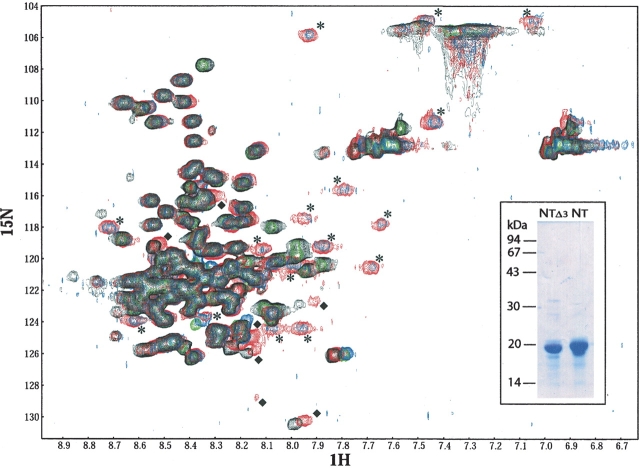
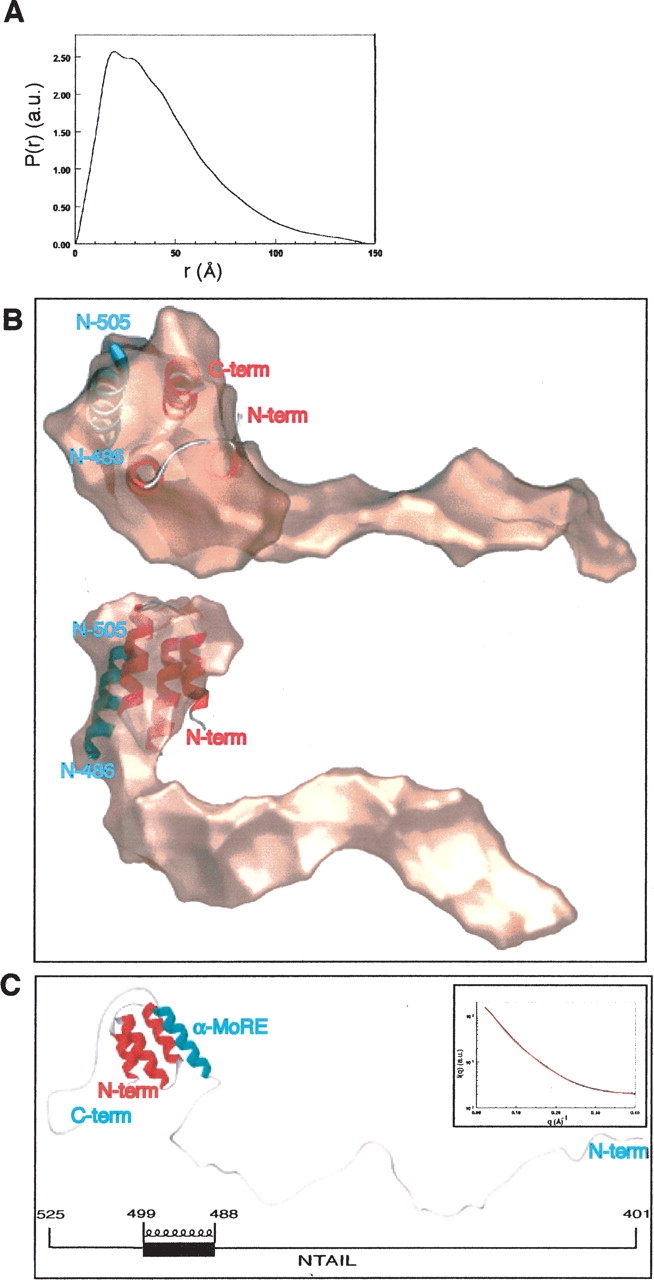
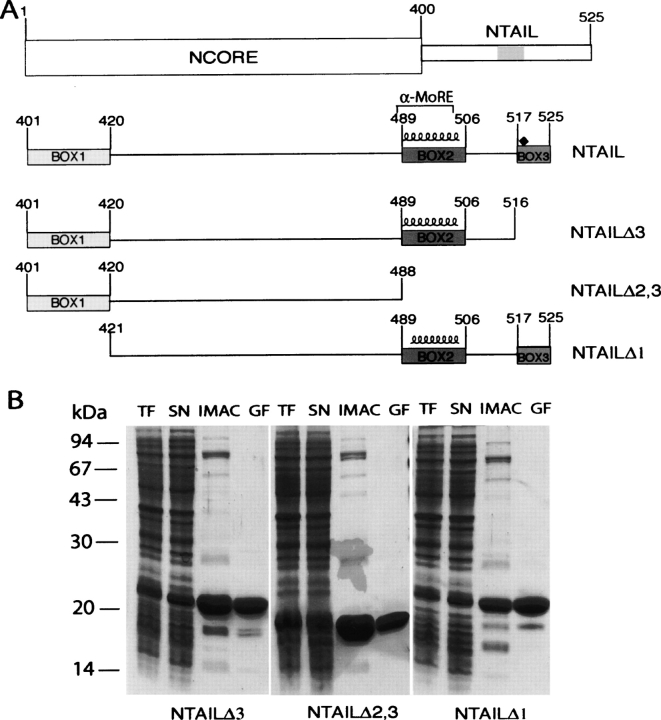
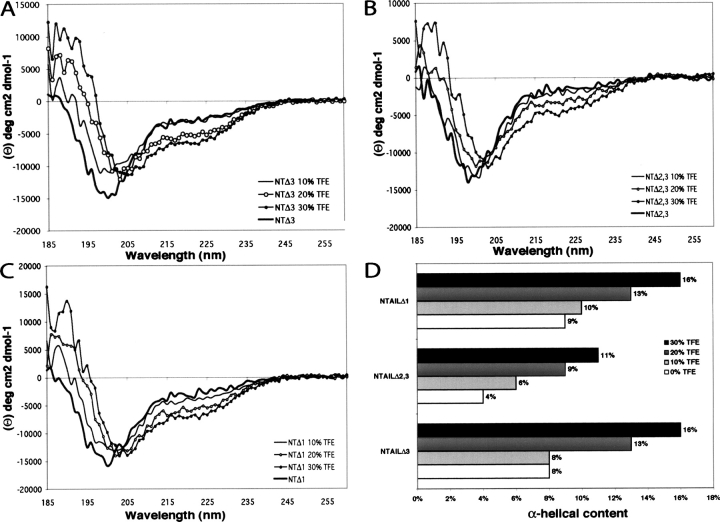
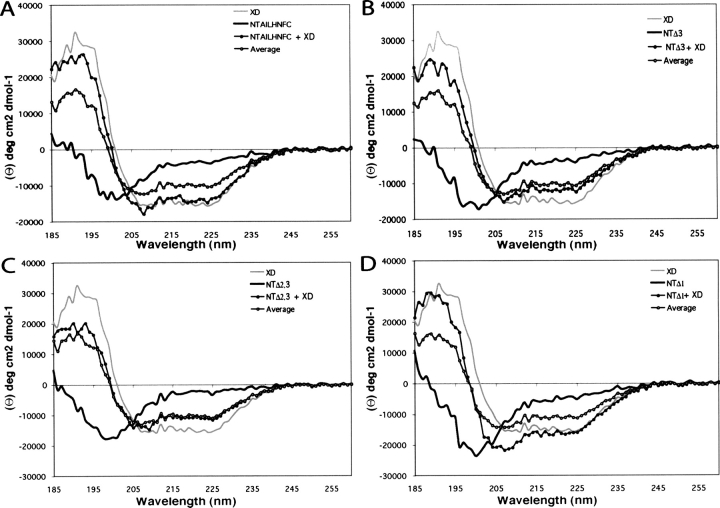
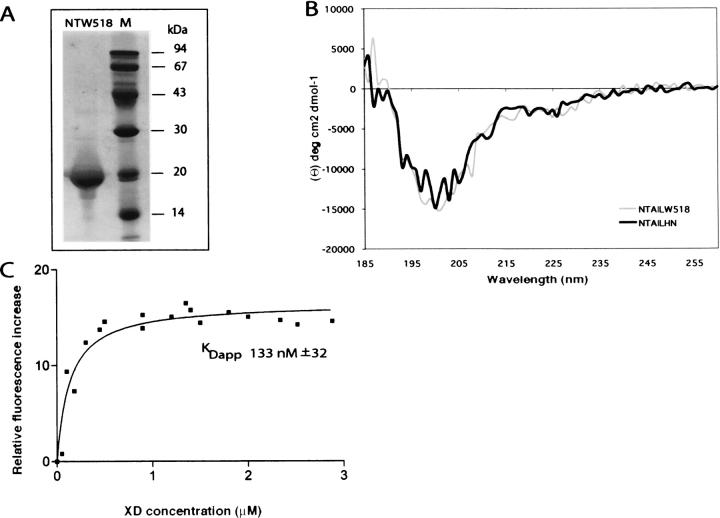
Measles virus is a negative-sense, single-stranded RNA virus within the Mononegavirales order,which includes several human pathogens, including rabies, Ebola, Nipah, and Hendra viruses. The measles virus nucleoprotein consists of a structured N-terminal domain, and of an intrinsically disordered C-terminal domain, N(TAIL) (aa 401-525), which undergoes induced folding in the presence of the C-terminal domain (XD, aa 459-507) of the viral phosphoprotein. With in N(TAIL), an alpha-helical molecular recognition element (alpha-MoRE, aa 488-499) involved in binding to P and in induced folding was identified and then observed in the crystal structure of XD. Using small-angle X-ray scattering, we have derived a low-resolution structural model of the complex between XD and N(TAIL), which shows that most of N(TAIL) remains disordered in the complex despite P-induced folding within the alpha-MoRE. The model consists of an extended shape accommodating the multiple conformations adopted by the disordered N-terminal region of N(TAIL), and of a bulky globular region, corresponding to XD and to the C terminus of N(TAIL) (aa 486-525). Using surface plasmon resonance, circular dichroism, fluorescence spectroscopy, and heteronuclear magnetic resonance, we show that N(TAIL) has an additional site (aa 517-525) involved in binding to XD but not in the unstructured-to-structured transition. This work provides evidence that intrinsically disordered domains can establish complex interactions with their partners, and can contact them through multiple sites that do not all necessarily gain regular secondary structure.
麻疹病毒是单股负链RNA病毒,属于单股负链RNA病毒目,该目还包括几种人类病原体,如狂犬病病毒、埃博拉病毒、尼帕病毒和亨德拉病毒。麻疹病毒核蛋白由一个结构化的N端结构域和一个内在无序的C端结构域N(TAIL)(氨基酸401 - 525)组成,在病毒磷蛋白的C端结构域(XD,氨基酸459 - 507)存在的情况下,N(TAIL)会发生诱导折叠。在N(TAIL)中,鉴定出了一个参与与P结合和诱导折叠的α-螺旋分子识别元件(α-MoRE,氨基酸488 - 499),随后在XD的晶体结构中观察到了该元件。利用小角X射线散射技术,我们推导了XD与N(TAIL)复合物的低分辨率结构模型,该模型表明,尽管α-MoRE内发生了P诱导的折叠,但复合物中N(TAIL)的大部分区域仍然无序。该模型由一个扩展形状组成,可容纳N(TAIL)无序N端区域所采用的多种构象,以及一个庞大的球状区域,对应于XD和N(TAIL)的C端(氨基酸486 - 525)。通过表面等离子体共振、圆二色性、荧光光谱和异核磁共振技术,我们发现N(TAIL)还有一个额外的结合位点(氨基酸517 - 525),该位点参与与XD的结合,但不参与非结构化到结构化的转变。这项工作提供了证据,证明内在无序结构域可以与其伙伴建立复杂的相互作用,并可以通过并非都一定会获得规则二级结构的多个位点与它们接触。