Agbottah Emmanuel, Deng Longwen, Dannenberg Luke O, Pumfery Anne, Kashanchi Fatah
The George Washington University Medical Center, Department of Biochemistry and Molecular Biology, Washington, DC 20037, USA.
Retrovirology. 2006 Aug 7;3:48. doi: 10.1186/1742-4690-3-48.
Human immunodeficiency virus type 1 (HIV-1) is the etiologic agent of acquired immunodeficiency virus (AIDS). Following entry into the host cell, the viral RNA is reverse transcribed into DNA and subsequently integrated into the host genome as a chromatin template. The integrated proviral DNA, along with the specific chromatinized environment in which integration takes place allows for the coordinated regulation of viral transcription and replication. While the specific roles of and interplay between viral and host proteins have not been fully elucidated, numerous reports indicate that HIV-1 retains the ability for self-regulation via the pleiotropic effects of its viral proteins. Though viral transcription is fully dependent upon host cellular factors and the state of host activation, recent findings indicate a complex interplay between viral proteins and host transcription regulatory machineries including histone deacetylases (HDACs), histone acetyltransferases (HATs), cyclin dependent kinases (CDKs), and histone methyltransferases (HMTs).
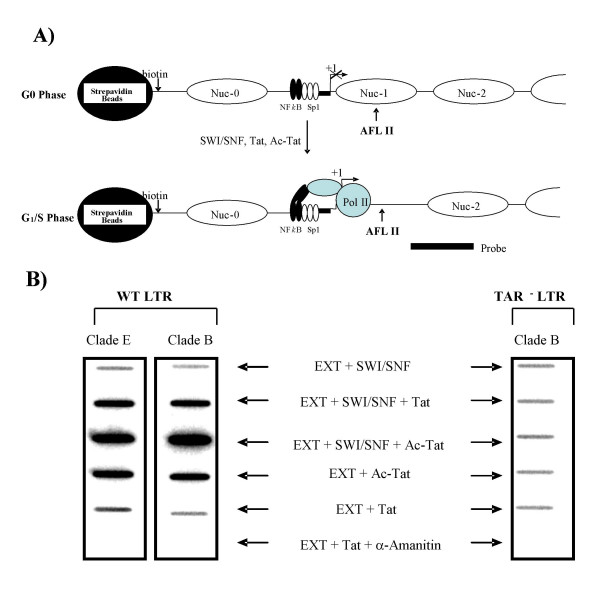
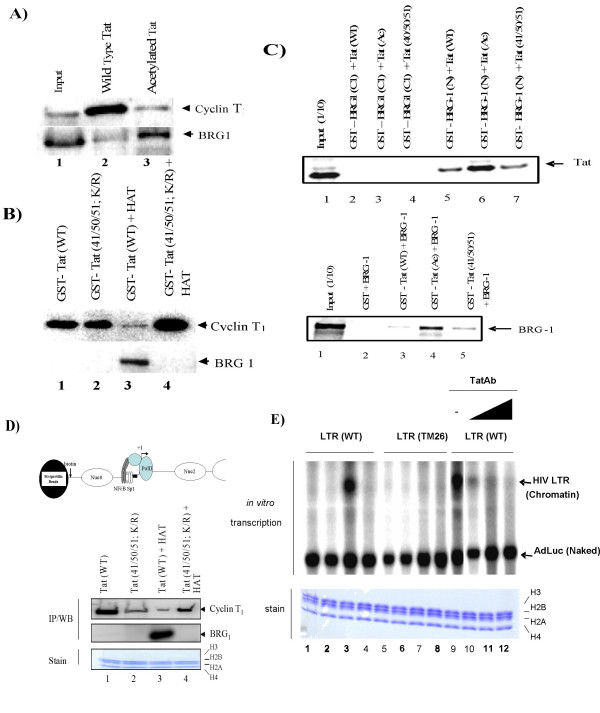
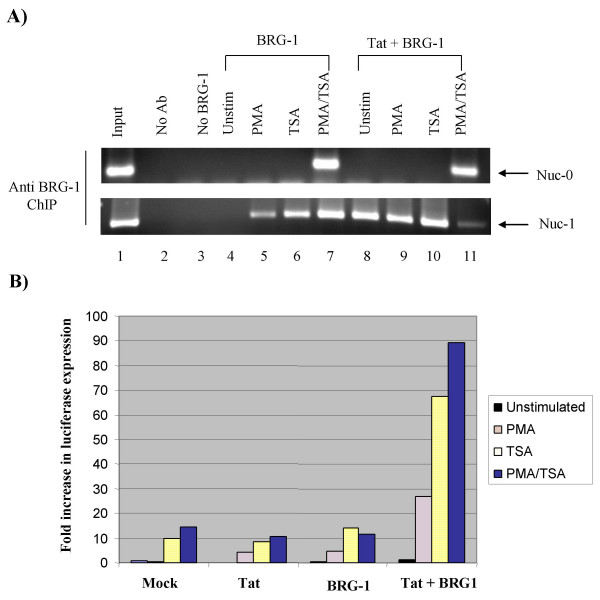
Here, we describe the effect of Tat activated transcription at the G1/S border of the cell cycle and analyze the interaction of modified Tat with the chromatin remodeling complex, SWI/SNF. HIV-1 LTR DNA reconstituted into nucleosomes can be activated in vitro using various Tat expressing extracts. Optimally activated transcription was observed at the G1/S border of the cell cycle both in vitro and in vivo, where chromatin remodeling complex, SWI/SNF, was present on the immobilized LTR DNA. Using a number of in vitro binding as well as in vivo chromatin immunoprecipitation (ChIP) assays, we detected the presence of both BRG1 and acetylated Tat in the same complex. Finally, we demonstrate that activated transcription resulted in partial or complete removal of the nucleosome from the start site of the LTR as evidenced by a restriction enzyme accessibility assay.
We propose a model where unmodified Tat is involved in binding to the CBP/p300 and cdk9/cyclin T1 complexes facilitating transcription initiation. Acetylated Tat dissociates from the TAR RNA structure and recruits bromodomain-binding chromatin modifying complexes such as p/CAF and SWI/SNF to possibly facilitate transcription elongation.
人类免疫缺陷病毒1型(HIV-1)是获得性免疫缺陷综合征(AIDS)的病原体。进入宿主细胞后,病毒RNA被逆转录为DNA,随后作为染色质模板整合到宿主基因组中。整合的前病毒DNA,连同整合发生时的特定染色质化环境,允许对病毒转录和复制进行协调调控。虽然病毒蛋白与宿主蛋白之间的具体作用及相互作用尚未完全阐明,但大量报告表明,HIV-1通过其病毒蛋白的多效性作用保留了自我调节能力。尽管病毒转录完全依赖于宿主细胞因子和宿主激活状态,但最近的研究结果表明,病毒蛋白与包括组蛋白去乙酰化酶(HDACs)、组蛋白乙酰转移酶(HATs)、细胞周期蛋白依赖性激酶(CDKs)和组蛋白甲基转移酶(HMTs)在内的宿主转录调节机制之间存在复杂的相互作用。
在此,我们描述了Tat在细胞周期的G1/S边界激活转录的作用,并分析了修饰的Tat与染色质重塑复合物SWI/SNF的相互作用。重组为核小体的HIV-1 LTR DNA可在体外使用各种表达Tat的提取物激活。在体外和体内的细胞周期G1/S边界均观察到最佳激活转录,其中染色质重塑复合物SWI/SNF存在于固定的LTR DNA上。使用多种体外结合以及体内染色质免疫沉淀(ChIP)试验,我们在同一复合物中检测到BRG1和乙酰化Tat的存在。最后,我们证明激活转录导致核小体从LTR的起始位点部分或完全去除,这通过限制性内切酶可及性试验得到证明。
我们提出了一个模型,其中未修饰的Tat参与与CBP/p300和cdk9/细胞周期蛋白T1复合物的结合,促进转录起始。乙酰化的Tat从TAR RNA结构解离,并募集诸如p/CAF和SWI/SNF等含溴结构域的染色质修饰复合物,可能促进转录延伸。