Schlesner Matthias, Miller Arthur, Streif Stefan, Staudinger Wilfried F, Müller Judith, Scheffer Beatrix, Siedler Frank, Oesterhelt Dieter
Department of Membrane Biochemistry, Max Planck Institute of Biochemistry, Am Klopferspitz 18, 82152 Martinsried, Germany.
BMC Microbiol. 2009 Mar 16;9:56. doi: 10.1186/1471-2180-9-56.
Archaea share with bacteria the ability to bias their movement towards more favorable locations, a process known as taxis. Two molecular systems drive this process: the motility apparatus and the chemotaxis signal transduction system. The first consists of the flagellum, the flagellar motor, and its switch, which allows cells to reverse the rotation of flagella. The second targets the flagellar motor switch in order to modulate the switching frequency in response to external stimuli. While the signal transduction system is conserved throughout archaea and bacteria, the archaeal flagellar apparatus is different from the bacterial one. The proteins constituting the flagellar motor and its switch in archaea have not yet been identified, and the connection between the bacterial-like chemotaxis signal transduction system and the archaeal motility apparatus is unknown.
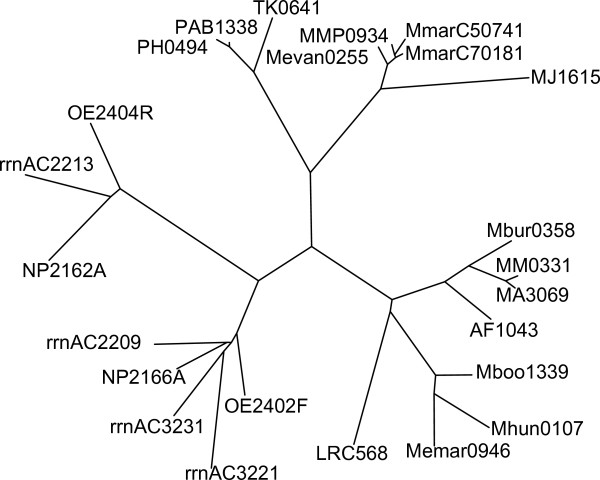
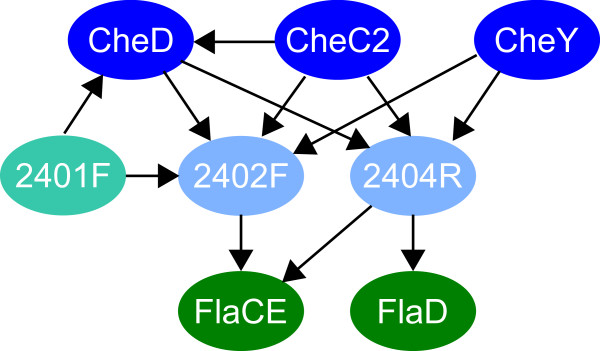
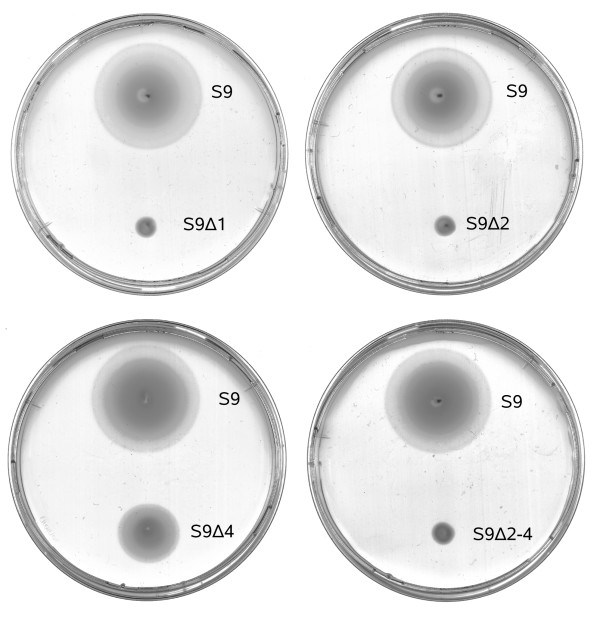
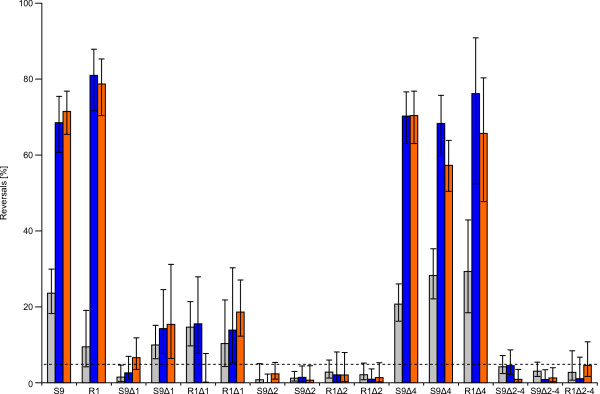
Using protein-protein interaction analysis, we have identified three proteins in Halobacterium salinarum that interact with the chemotaxis (Che) proteins CheY, CheD, and CheC2, as well as the flagella accessory (Fla) proteins FlaCE and FlaD. Two of the proteins belong to the protein family DUF439, the third is a HEAT_PBS family protein. In-frame deletion strains for all three proteins were generated and analyzed as follows: a) photophobic responses were measured by a computer-based cell tracking system b) flagellar rotational bias was determined by dark-field microscopy, and c) chemotactic behavior was analyzed by a swarm plate assay. Strains deleted for the HEAT_PBS protein or one of the DUF439 proteins proved unable to switch the direction of flagellar rotation. In these mutants, flagella rotate only clockwise, resulting in exclusively forward swimming cells that are unable to respond to tactic signals. Deletion of the second DUF439 protein had only minimal effects. HEAT_PBS proteins could be identified in the chemotaxis gene regions of all motile haloarchaea sequenced so far, but not in those of other archaeal species. Genes coding for DUF439 proteins, however, were found to be integral parts of chemotaxis gene regions across the archaeal domain, and they were not detected in other genomic context.
Altogether, these results demonstrate that, in the archaeal domain, previously unrecognized archaea-specific Che proteins are essential for relaying taxis signaling to the flagellar apparatus.
古菌与细菌一样,具有使自身朝着更适宜的位置移动的能力,这一过程称为趋化性。有两个分子系统驱动这一过程:运动装置和趋化信号转导系统。第一个系统由鞭毛、鞭毛马达及其开关组成,该开关可使细胞逆转鞭毛的旋转方向。第二个系统作用于鞭毛马达开关,以便根据外部刺激调节开关频率。虽然信号转导系统在整个古菌和细菌中是保守的,但古菌的鞭毛装置与细菌的不同。构成古菌鞭毛马达及其开关的蛋白质尚未被鉴定出来,类似细菌的趋化信号转导系统与古菌运动装置之间的联系也不清楚。
通过蛋白质-蛋白质相互作用分析,我们在盐生盐杆菌中鉴定出三种蛋白质,它们与趋化蛋白CheY、CheD和CheC2以及鞭毛辅助蛋白FlaCE和FlaD相互作用。其中两种蛋白质属于DUF439蛋白家族,第三种是HEAT_PBS家族蛋白。构建了这三种蛋白质的框内缺失菌株并进行如下分析:a)通过基于计算机的细胞追踪系统测量避光反应;b)通过暗场显微镜确定鞭毛旋转偏向;c)通过群体平板试验分析趋化行为。缺失HEAT_PBS蛋白或其中一种DUF439蛋白的菌株无法改变鞭毛旋转方向。在这些突变体中,鞭毛仅顺时针旋转,导致细胞只能向前游动,无法对趋化信号作出反应。缺失第二种DUF439蛋白的影响很小。到目前为止,在所有已测序的运动型嗜盐古菌的趋化基因区域中都能鉴定出HEAT_PBS蛋白,但在其他古菌物种中则没有。然而,编码DUF439蛋白的基因是古菌域趋化基因区域的组成部分,在其他基因组背景中未检测到。
总之,这些结果表明,在古菌域中,以前未被识别的古菌特异性趋化蛋白对于将趋化信号传递到鞭毛装置至关重要。