Arya Gaurav, Schlick Tamar
Department of Nanoengineering, University of California, San Diego, 9500 Gilman Drive, Mail Code: 0448, La Jolla, California 92093, USA.
J Phys Chem A. 2009 Apr 23;113(16):4045-59. doi: 10.1021/jp810375d.
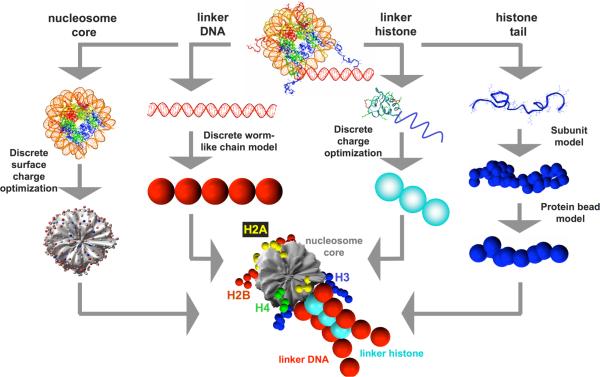
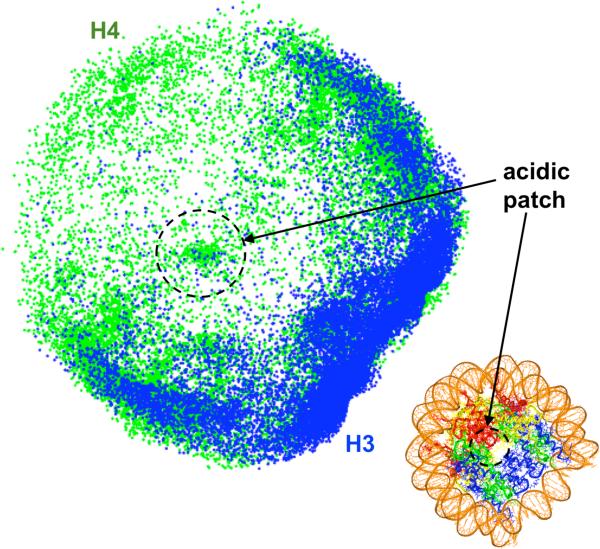
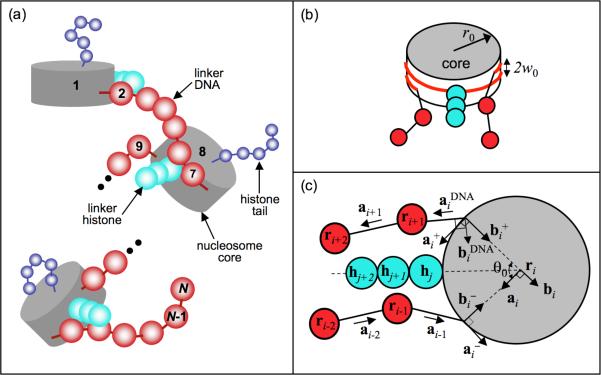
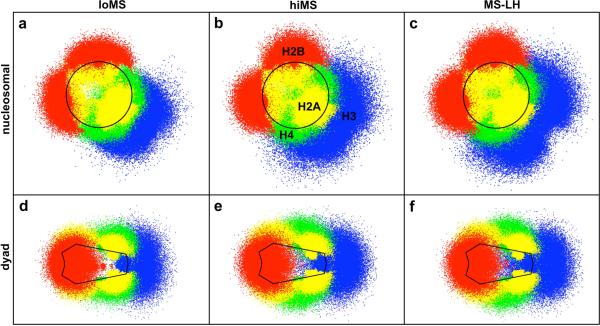
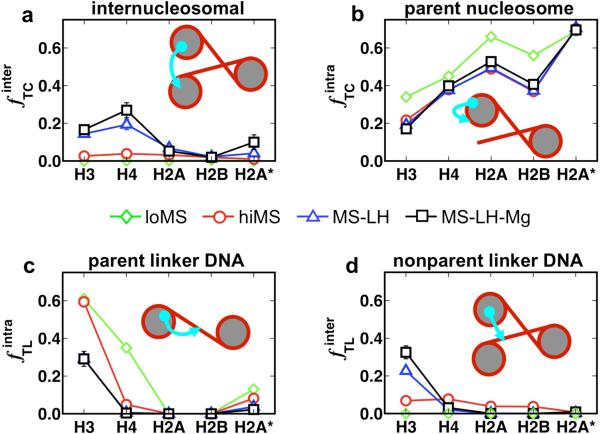
To elucidate the role of the histone tails in chromatin compaction and in higher-order folding of chromatin under physiological conditions, we extend a mesoscale model of chromatin (Arya, Zhang, and Schlick. Biophys. J. 2006, 91, 133; Arya and Schlick. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 16236) to account for divalent cations (Mg(2+)) and linker histones. Configurations of 24-nucleosome oligonucleosomes in different salt environments and in the presence and absence of linker histones are sampled by a mixture of local and global Monte Carlo methods. Analyses of the resulting ensembles reveal a dynamic synergism between the histone tails, linker histones, and ions in forming compact higher-order structures of chromatin. In the presence of monovalent salt alone, oligonucleosomes remain relatively unfolded, and the histone tails do not mediate many internucleosomal interactions. Upon the addition of linker histones and divalent cations, the oligonucleosomes undergo a significant compaction triggered by a dramatic increase in the internucleosomal interactions mediated by the histone tails, formation of a rigid linker DNA "stem" around the linker histones' C-terminal domains, and reduction in the electrostatic repulsion between linker DNAs via sharp bending in some linker DNAs caused by the divalent cations. Among all histone tails, the H4 tails mediate the most internucleosomal interactions, consistent with experimental observations, followed by the H3, H2A, and H2B tails in decreasing order. Apart from mediating internucleosomal interactions, the H3 tails also contribute to chromatin compaction by attaching to the entering and exiting linker DNA to screen electrotatic repulsion among the linker DNAs. This tendency of the H3 tails to attach to linker DNA, however, decreases significantly upon the addition of linker histones due to competition effects. The H2A and H2B tails do not mediate significant internucleosomal interactions but are important for mediating fiber/fiber intractions, especially in relatively unfolded chromatin in monovalent salt environments.
为了阐明组蛋白尾部在生理条件下染色质压缩和染色质高级折叠中的作用,我们扩展了一个染色质的中尺度模型(Arya、Zhang和Schlick,《生物物理杂志》,2006年,91卷,133页;Arya和Schlick,《美国国家科学院院刊》,2006年,103卷,16236页),以考虑二价阳离子(Mg(2+))和连接组蛋白。通过局部和全局蒙特卡罗方法的混合,对不同盐环境下以及存在和不存在连接组蛋白时24核小体寡核小体的构型进行采样。对所得集合的分析揭示了组蛋白尾部、连接组蛋白和离子在形成染色质紧密高级结构中的动态协同作用。仅存在单价盐时,寡核小体保持相对伸展状态,组蛋白尾部不介导许多核小体间的相互作用。加入连接组蛋白和二价阳离子后,寡核小体经历显著压缩,这是由组蛋白尾部介导的核小体间相互作用急剧增加、连接组蛋白C末端结构域周围形成刚性连接DNA“茎”以及二价阳离子导致一些连接DNA急剧弯曲从而减少连接DNA之间的静电排斥所触发的。在所有组蛋白尾部中,H4尾部介导的核小体间相互作用最多,这与实验观察结果一致,其次是H3、H2A和H2B尾部,顺序递减。除了介导核小体间相互作用外,H3尾部还通过附着于进入和离开的连接DNA来屏蔽连接DNA之间的静电排斥,从而有助于染色质压缩。然而,由于竞争效应,加入连接组蛋白后,H3尾部附着于连接DNA的这种趋势显著降低。H2A和H2B尾部不介导显著的核小体间相互作用,但对于介导纤维/纤维相互作用很重要,特别是在单价盐环境中相对伸展的染色质中。