Department of Pharmacology, The School of Pharmacy, Brunswick Square, London, UK.
Hum Mol Genet. 2009 Oct 15;18(20):3955-68. doi: 10.1093/hmg/ddp337. Epub 2009 Jul 22.
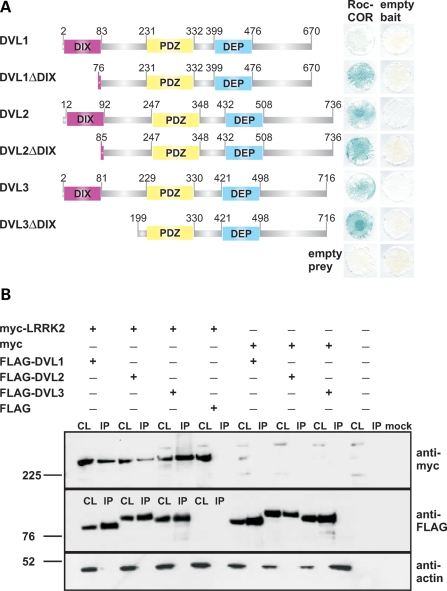
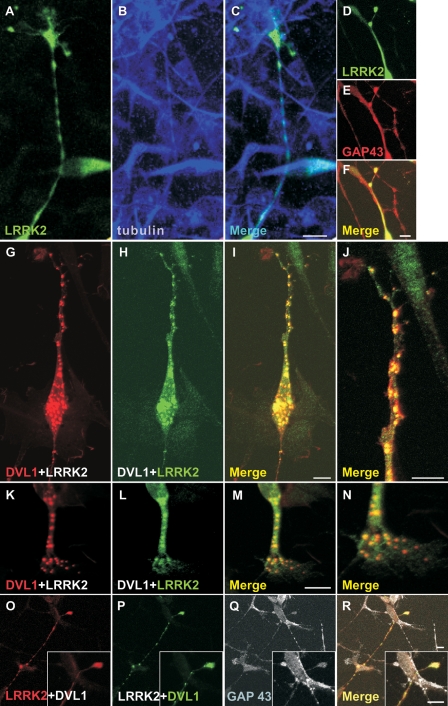
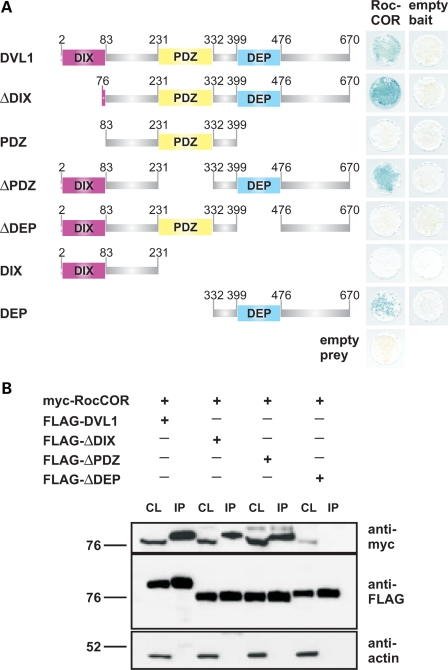
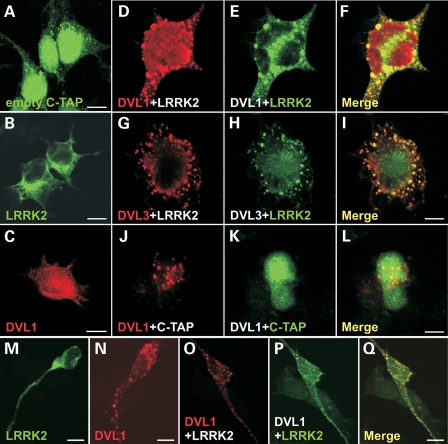
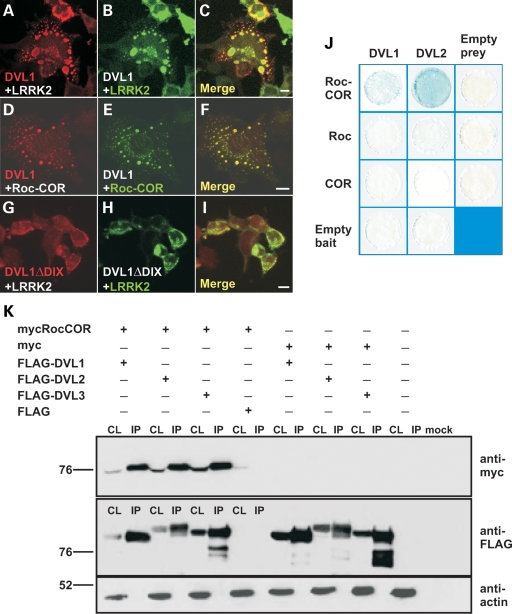
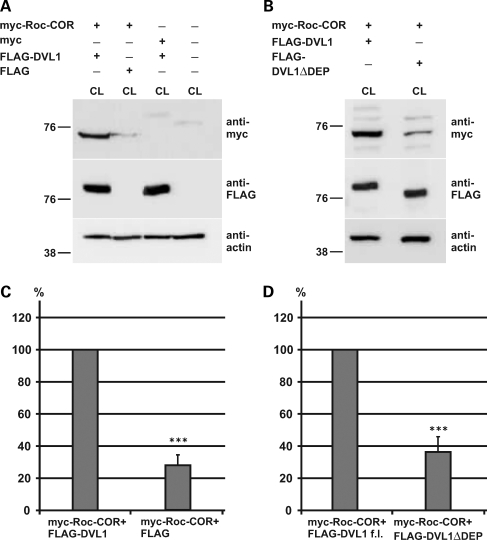
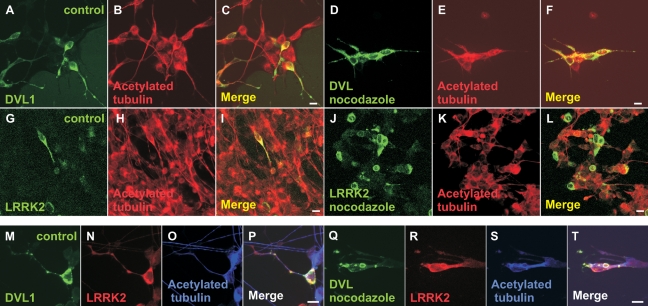
Mutations in PARK8, encoding LRRK2, are the most common known cause of Parkinson's disease. The LRRK2 Roc-COR tandem domain exhibits GTPase activity controlling LRRK2 kinase activity via an intramolecular process. We report the interaction of LRRK2 with the dishevelled family of phosphoproteins (DVL1-3), key regulators of Wnt (Wingless/Int) signalling pathways important for axon guidance, synapse formation and neuronal maintenance. Interestingly, DVLs can interact with and mediate the activation of small GTPases with structural similarity to the LRRK2 Roc domain. The LRRK2 Roc-COR domain and the DVL1 DEP domain were necessary and sufficient for LRRK2-DVL1 interaction. Co-expression of DVL1 increased LRRK2 steady-state protein levels, an effect that was dependent on the DEP domain. Strikingly, LRRK2-DVL1-3 interactions were disrupted by the familial PARK8 mutation Y1699C, whereas pathogenic mutations at residues R1441 and R1728 strengthened LRRK2-DVL1 interactions. Co-expression of DVL1 with LRRK2 in mammalian cells resulted in the redistribution of LRRK2 to typical cytoplasmic DVL1 aggregates in HEK293 and SH-SY5Y cells and co-localization in neurites and growth cones of differentiated dopaminergic SH-SY5Y cells. This is the first report of the modulation of a key LRRK2-accessory protein interaction by PARK8 Roc-COR domain mutations segregating with Parkinson's disease. Since the DVL1 DEP domain is known to be involved in the regulation of small GTPases, we propose that: (i) DVLs may influence LRRK2 GTPase activity, and (ii) Roc-COR domain mutations modulating LRRK2-DVL interactions indirectly influence kinase activity. Our findings also link LRRK2 to Wnt signalling pathways, suggesting novel pathogenic mechanisms and new targets for genetic analysis in Parkinson's disease.
LRRK2 基因突变是帕金森病最常见的已知原因。LRRK2 的 Roc-COR 串联结构域具有 GTPase 活性,通过分子内过程控制 LRRK2 激酶活性。我们报告了 LRRK2 与 DVL1-3 蛋白家族的相互作用,DVL1-3 蛋白家族是 Wnt(无翅型/Int)信号通路的关键调节因子,该信号通路对轴突导向、突触形成和神经元维持很重要。有趣的是,DVLs 可以与具有与 LRRK2 Roc 结构域相似结构的小 GTPase 相互作用并介导其激活。LRRK2 的 Roc-COR 结构域和 DVL1 的 DEP 结构域是 LRRK2-DVL1 相互作用的必要和充分条件。DVL1 的共表达增加了 LRRK2 的稳态蛋白水平,这一效应依赖于 DEP 结构域。令人惊讶的是,家族性 PARK8 突变 Y1699C 破坏了 LRRK2-DVL1-3 的相互作用,而残基 R1441 和 R1728 的致病性突变则增强了 LRRK2-DVL1 的相互作用。在哺乳动物细胞中共表达 DVL1 和 LRRK2 导致 LRRK2 在 HEK293 和 SH-SY5Y 细胞中重新分布到典型的细胞质 DVL1 聚集体中,并在分化的多巴胺能 SH-SY5Y 细胞的轴突和生长锥中发生共定位。这是首次报道 PARK8 Roc-COR 结构域突变调节与帕金森病相关的关键 LRRK2 辅助蛋白相互作用。由于 DVL1 的 DEP 结构域已知参与小 GTPase 的调节,我们提出:(i)DVLs 可能影响 LRRK2 GTPase 活性,(ii)调节 LRRK2-DVL 相互作用的 Roc-COR 结构域突变间接影响激酶活性。我们的研究结果还将 LRRK2 与 Wnt 信号通路联系起来,表明帕金森病中有新的致病机制和新的遗传分析靶点。