Laboratory of Molecular Neurobiology, Department of Medical Biochemistry and Biophysics, Karolinska Institutet, 17177, Stockholm, Sweden.
Current address: Science for Life Laboratory, Department of Biophysics and Biochemistry, Stockholm University, 171 65, Stockholm, Sweden.
Mol Neurodegener. 2017 Jul 11;12(1):54. doi: 10.1186/s13024-017-0193-9.
Autosomal-dominant mutations in the Park8 gene encoding Leucine-rich repeat kinase 2 (LRRK2) have been identified to cause up to 40% of the genetic forms of Parkinson's disease. However, the function and molecular pathways regulated by LRRK2 are largely unknown. It has been shown that LRRK2 serves as a scaffold during activation of WNT/β-catenin signaling via its interaction with the β-catenin destruction complex, DVL1-3 and LRP6. In this study, we examine whether LRRK2 also interacts with signaling components of the WNT/Planar Cell Polarity (WNT/PCP) pathway, which controls the maturation of substantia nigra dopaminergic neurons, the main cell type lost in Parkinson's disease patients.
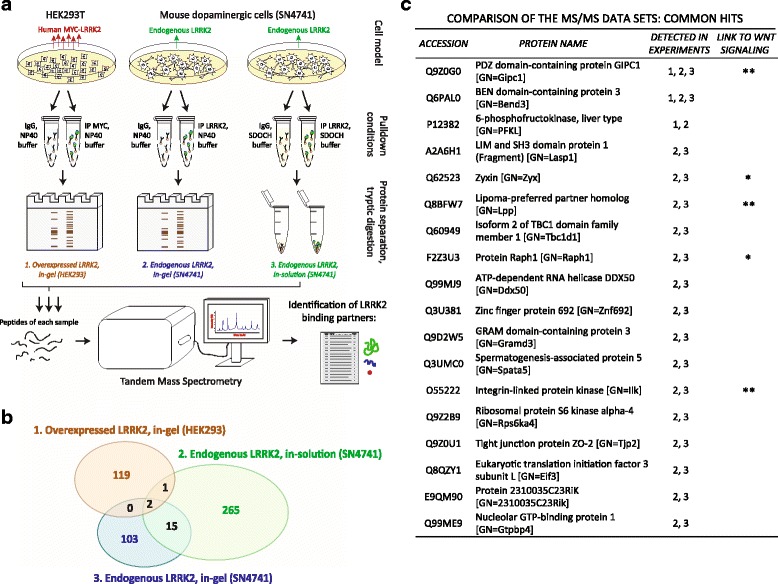
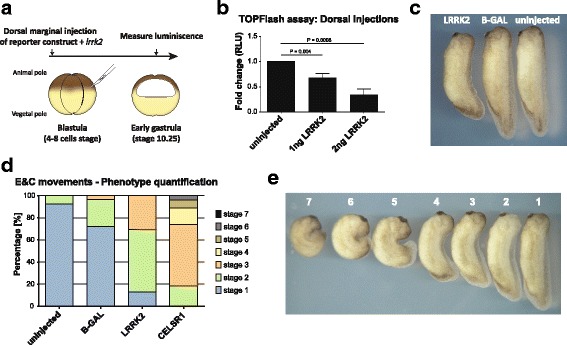
Co-immunoprecipitation and tandem mass spectrometry was performed in a mouse substantia nigra cell line (SN4741) and human HEK293T cell line in order to identify novel LRRK2 binding partners. Inhibition of the WNT/β-catenin reporter, TOPFlash, was used as a read-out of WNT/PCP pathway activation. The capacity of LRRK2 to regulate WNT/PCP signaling in vivo was tested in Xenopus laevis' early development.
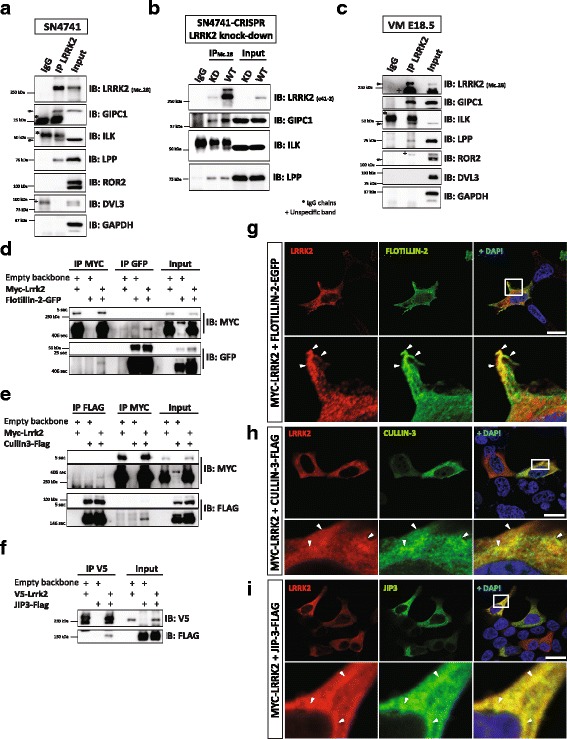
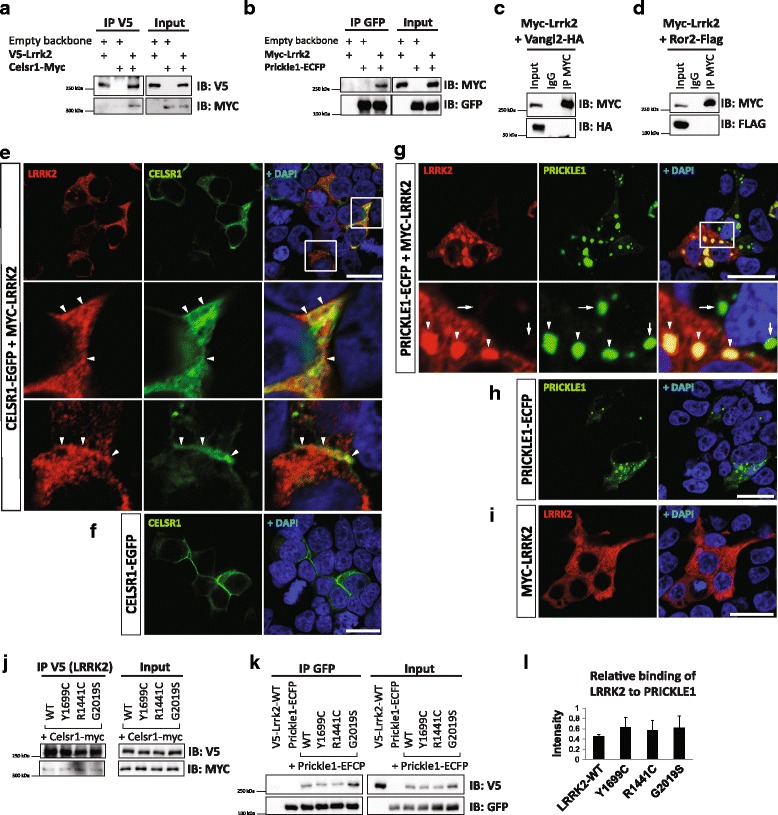
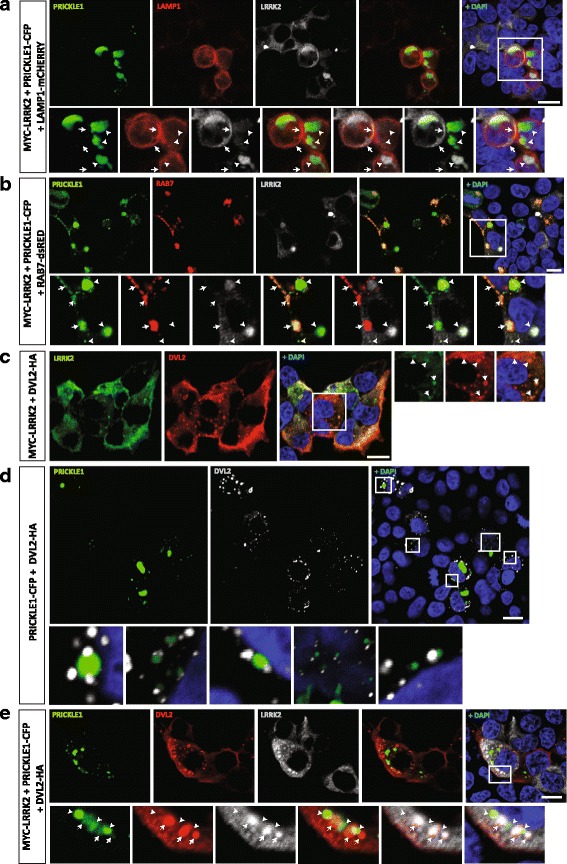
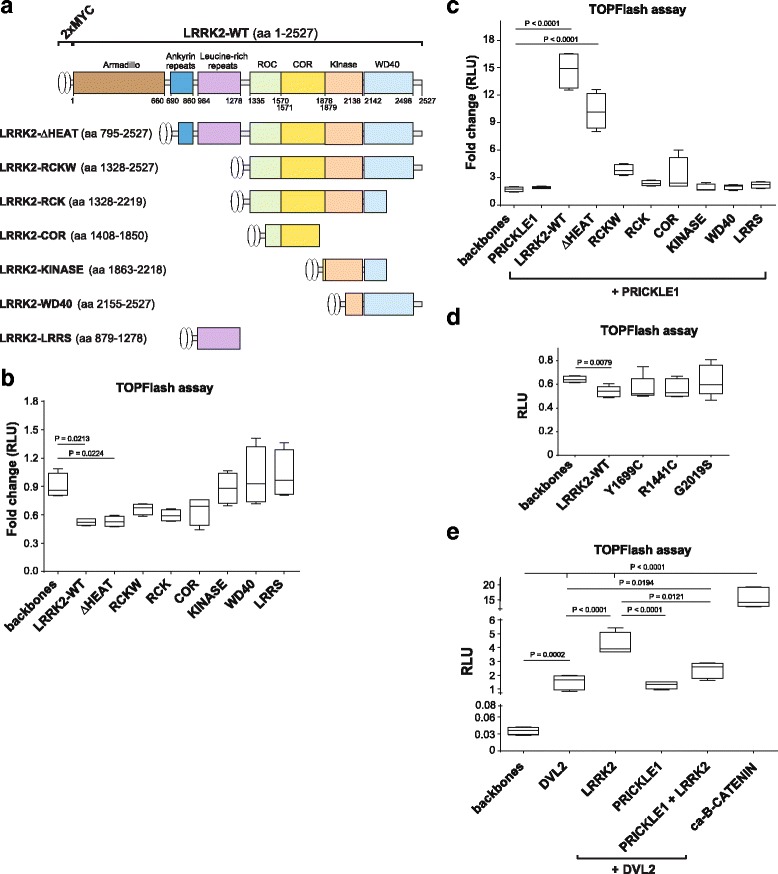
Our proteomic analysis identified that LRRK2 interacts with proteins involved in WNT/PCP signaling such as the PDZ domain-containing protein GIPC1 and Integrin-linked kinase (ILK) in dopaminergic cells in vitro and in the mouse ventral midbrain in vivo. Moreover, co-immunoprecipitation analysis revealed that LRRK2 binds to two core components of the WNT/PCP signaling pathway, PRICKLE1 and CELSR1, as well as to FLOTILLIN-2 and CULLIN-3, which regulate WNT secretion and inhibit WNT/β-catenin signaling, respectively. We also found that PRICKLE1 and LRRK2 localize in signalosomes and act as dual regulators of WNT/PCP and β-catenin signaling. Accordingly, analysis of the function of LRRK2 in vivo, in X. laevis revelaed that LRKK2 not only inhibits WNT/β-catenin pathway, but induces a classical WNT/PCP phenotype in vivo.
Our study shows for the first time that LRRK2 activates the WNT/PCP signaling pathway through its interaction to multiple WNT/PCP components. We suggest that LRRK2 regulates the balance between WNT/β-catenin and WNT/PCP signaling, depending on the binding partners. Since this balance is crucial for homeostasis of midbrain dopaminergic neurons, we hypothesize that its alteration may contribute to the pathophysiology of Parkinson's disease.
导致高达 40%的遗传形式帕金森病的原因是编码富亮氨酸重复激酶 2(LRRK2)的常染色体显性突变。然而,LRRK2 调节的功能和分子途径在很大程度上尚不清楚。已经表明,LRRK2 通过与β-连环蛋白破坏复合物、DVL1-3 和 LRP6 的相互作用,作为 WNT/β-连环蛋白信号传导的支架。在这项研究中,我们检查了 LRRK2 是否也与 WNT/平面细胞极性(WNT/PCP)途径的信号成分相互作用,该途径控制黑质多巴胺能神经元的成熟,这是帕金森病患者中丢失的主要细胞类型。
为了鉴定新的 LRRK2 结合伙伴,在鼠黑质细胞系(SN4741)和人 HEK293T 细胞系中进行了免疫沉淀和串联质谱分析。WNT/PCP 途径激活的 WNT/β-连环蛋白报告物 TOPFlash 的抑制作用用作 WNT/PCP 信号传导的读出。在非洲爪蟾早期发育中测试了 LRRK2 调节体内 WNT/PCP 信号传导的能力。
我们的蛋白质组学分析表明,LRRK2 在体外和体内的多巴胺能细胞中与 PDZ 结构域蛋白 GIPC1 和整合素连接激酶(ILK)等参与 WNT/PCP 信号传导的蛋白相互作用。此外,免疫沉淀分析显示 LRRK2 与 WNT/PCP 信号通路的两个核心成分 PRICKLE1 和 CELSR1 以及调节 WNT 分泌和抑制 WNT/β-连环蛋白信号传导的 FLOTILLIN-2 和 CULLIN-3 结合。我们还发现 PRICKLE1 和 LRRK2 定位于信号小体中,并作为 WNT/PCP 和 β-连环蛋白信号传导的双重调节剂。因此,对非洲爪蟾体内 LRRK2 功能的分析表明,LRRK2 不仅抑制 WNT/β-连环蛋白途径,而且在体内诱导经典的 WNT/PCP 表型。
我们的研究首次表明,LRRK2 通过与多个 WNT/PCP 成分相互作用来激活 WNT/PCP 信号通路。我们认为,LRRK2 根据结合伙伴调节 WNT/β-连环蛋白和 WNT/PCP 信号之间的平衡。由于这种平衡对于中脑多巴胺能神经元的内稳态至关重要,我们假设其改变可能导致帕金森病的病理生理学。