Institute of Physiological Chemistry, Technische Universität Dresden, Dresden, Germany.
PLoS Genet. 2009 Oct;5(10):e1000702. doi: 10.1371/journal.pgen.1000702. Epub 2009 Oct 23.
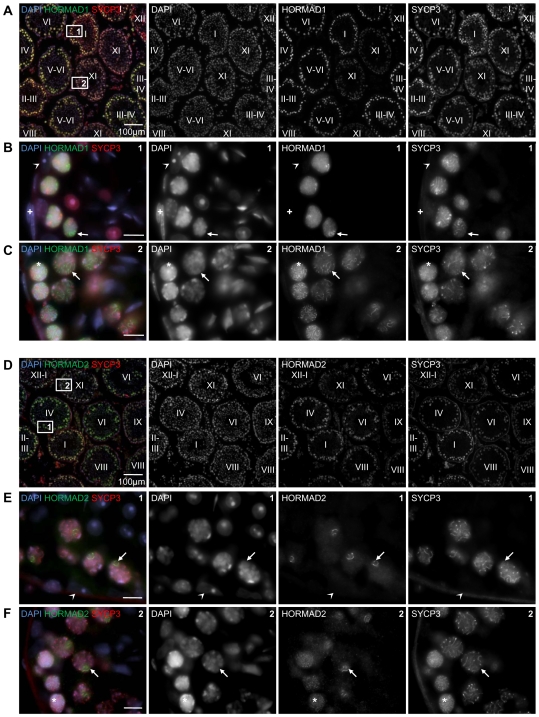
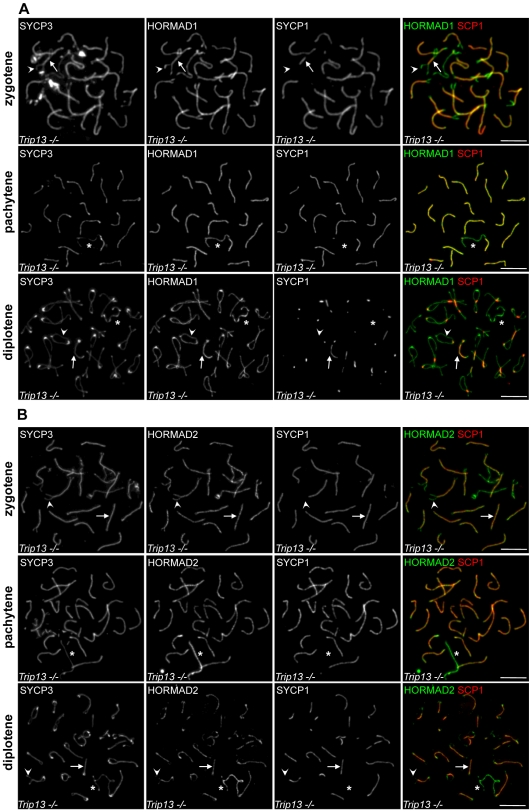
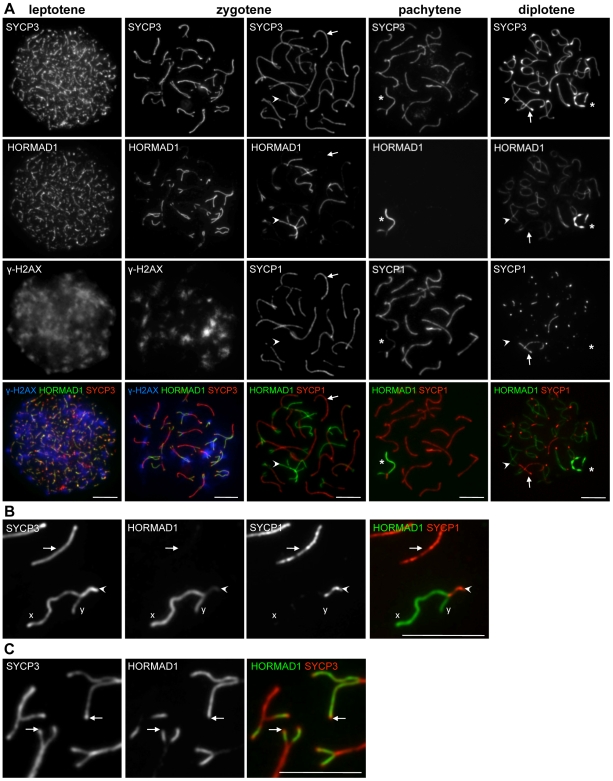
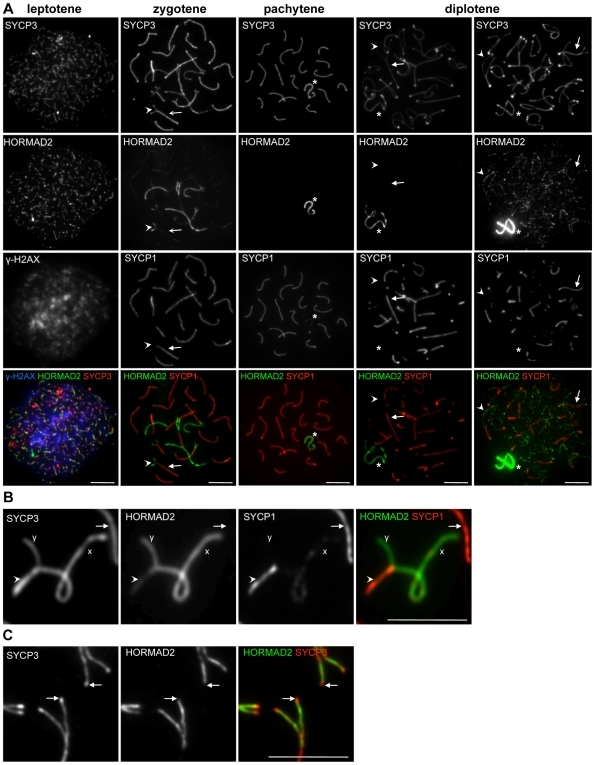
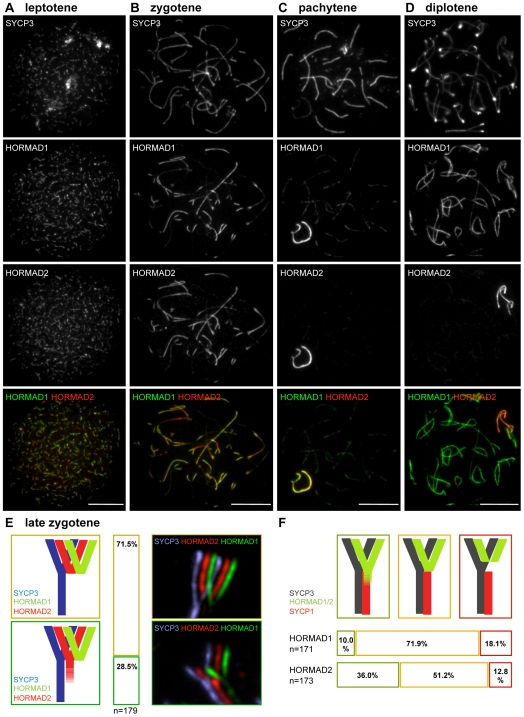
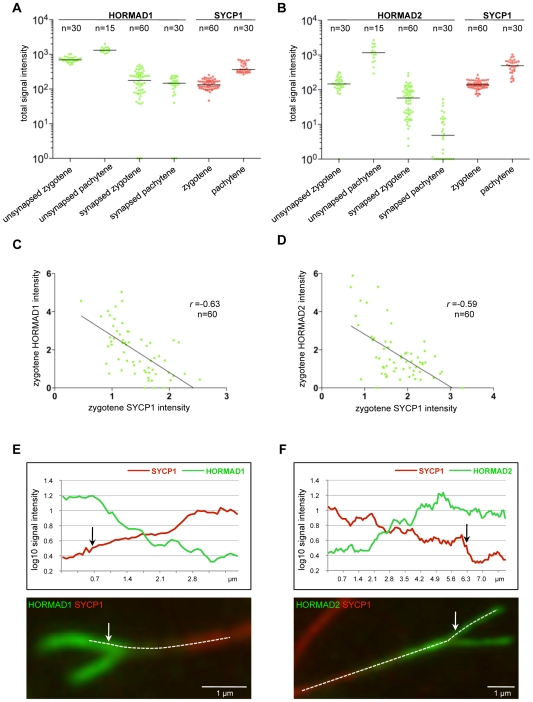
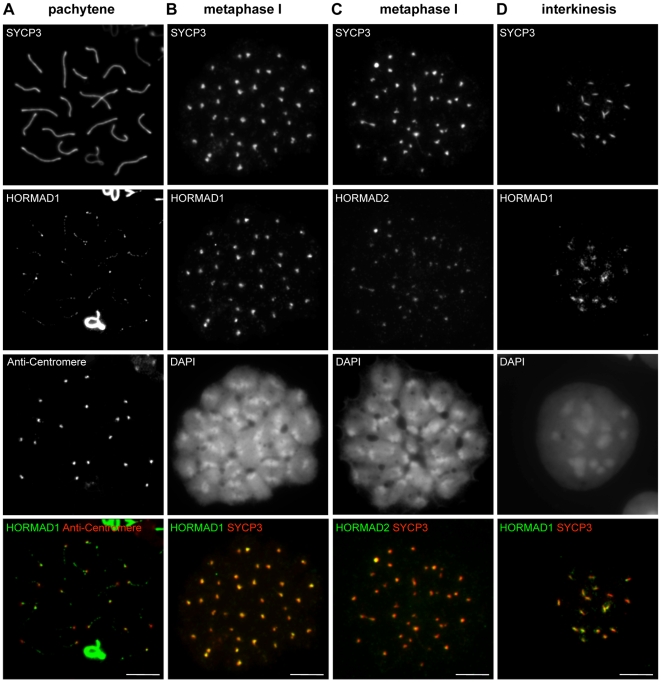
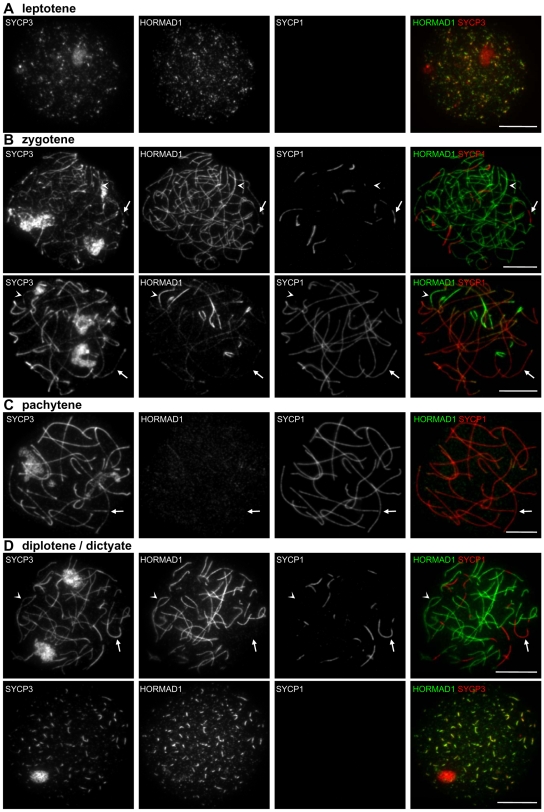
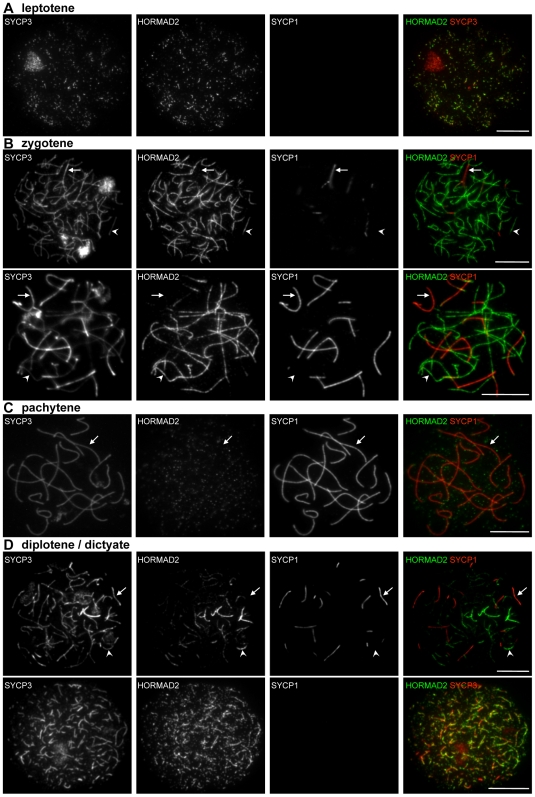
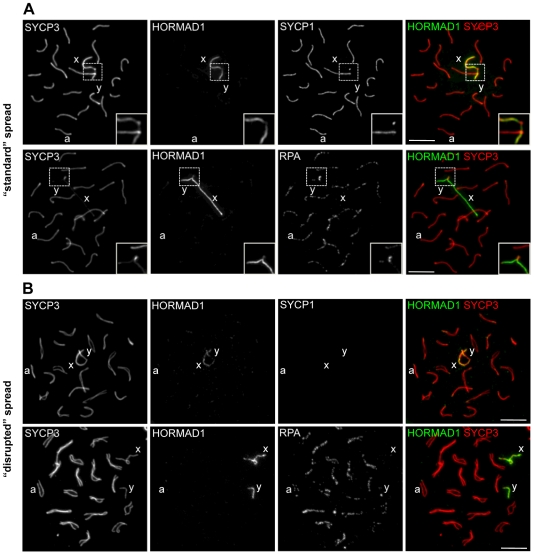
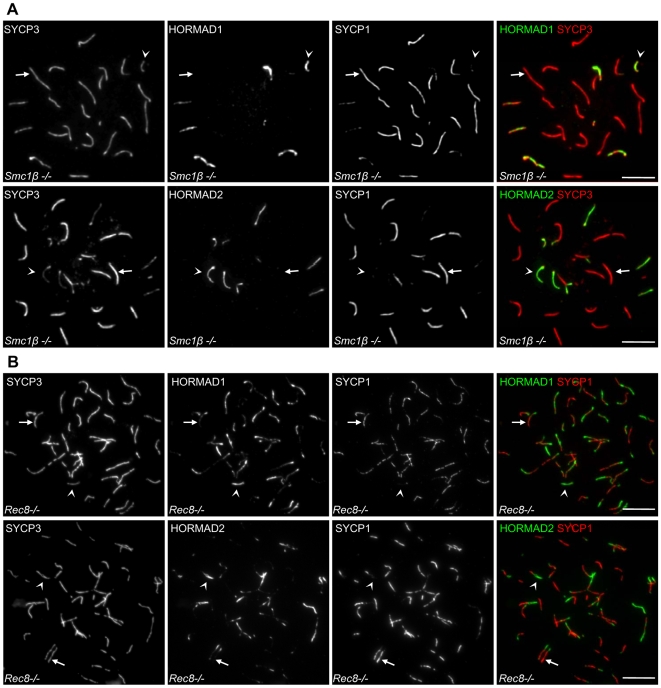
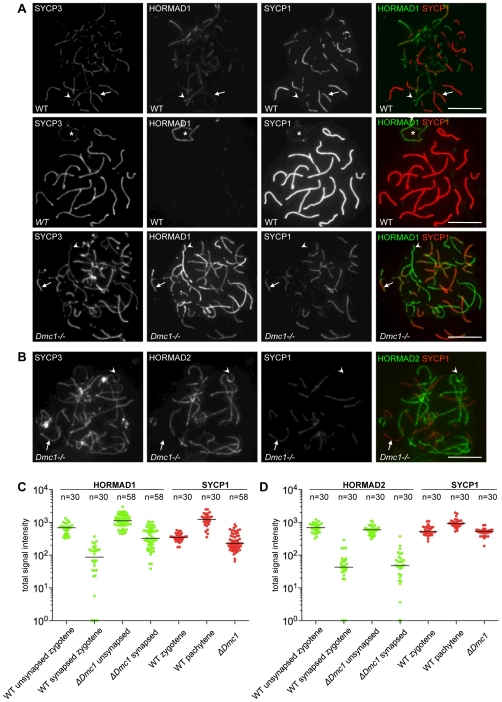
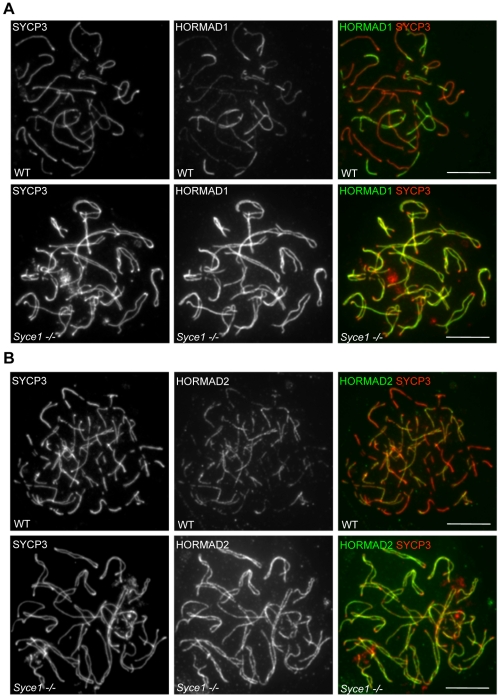
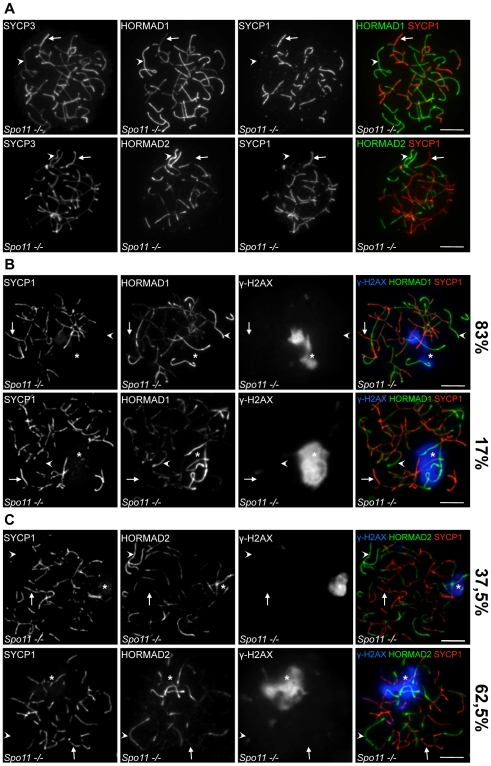
Meiotic crossovers are produced when programmed double-strand breaks (DSBs) are repaired by recombination from homologous chromosomes (homologues). In a wide variety of organisms, meiotic HORMA-domain proteins are required to direct DSB repair towards homologues. This inter-homologue bias is required for efficient homology search, homologue alignment, and crossover formation. HORMA-domain proteins are also implicated in other processes related to crossover formation, including DSB formation, inhibition of promiscuous formation of the synaptonemal complex (SC), and the meiotic prophase checkpoint that monitors both DSB processing and SCs. We examined the behavior of two previously uncharacterized meiosis-specific mouse HORMA-domain proteins--HORMAD1 and HORMAD2--in wild-type mice and in mutants defective in DSB processing or SC formation. HORMADs are preferentially associated with unsynapsed chromosome axes throughout meiotic prophase. We observe a strong negative correlation between SC formation and presence of HORMADs on axes, and a positive correlation between the presumptive sites of high checkpoint-kinase ATR activity and hyper-accumulation of HORMADs on axes. HORMADs are not depleted from chromosomes in mutants that lack SCs. In contrast, DSB formation and DSB repair are not absolutely required for depletion of HORMADs from synapsed axes. A simple interpretation of these findings is that SC formation directly or indirectly promotes depletion of HORMADs from chromosome axes. We also find that TRIP13 protein is required for reciprocal distribution of HORMADs and the SYCP1/SC-component along chromosome axes. Similarities in mouse and budding yeast meiosis suggest that TRIP13/Pch2 proteins have a conserved role in establishing mutually exclusive HORMAD-rich and synapsed chromatin domains in both mouse and yeast. Taken together, our observations raise the possibility that involvement of meiotic HORMA-domain proteins in the regulation of homologue interactions is conserved in mammals.
减数分裂交叉是由同源染色体(同源物)的重组修复程序性双链断裂(DSB)产生的。在各种各样的生物体中,减数分裂 HORMA 结构域蛋白需要指导 DSB 修复到同源物。这种同源物之间的偏向是有效同源搜索、同源物对齐和交叉形成所必需的。HORMA 结构域蛋白也与其他与交叉形成相关的过程有关,包括 DSB 的形成、抑制联会复合体(SC)的乱交形成,以及监测 DSB 加工和 SC 的减数分裂前期检查点。我们研究了两种以前未表征的减数分裂特异性小鼠 HORMA 结构域蛋白-HORMAD1 和 HORMAD2-在野生型小鼠和 DSB 加工或 SC 形成缺陷的突变体中的行为。HORMADs 在减数分裂前期的整个过程中优先与未联会的染色体轴相关联。我们观察到 SC 形成与轴上 HORMADs 的存在之间存在强烈的负相关,以及假定的高检查点激酶 ATR 活性部位与轴上 HORMADs 的过度积累之间存在正相关。在缺乏 SC 的突变体中,HORMADs 不会从染色体中耗尽。相反,DSB 的形成和修复对于 HORMADs 从联会轴中耗尽并非绝对必要。对这些发现的一个简单解释是,SC 的形成直接或间接地促进了 HORMADs 从染色体轴上的耗尽。我们还发现 TRIP13 蛋白对于 HORMAD 和 SYCP1/SC 成分沿着染色体轴的相互分布是必需的。在小鼠和芽殖酵母减数分裂中存在相似性表明,TRIP13/Pch2 蛋白在建立这两种生物中互斥的富含 HORMAD 和联会染色质域方面具有保守作用。总之,我们的观察结果提出了这样一种可能性,即减数分裂 HORMA 结构域蛋白在同源物相互作用的调节中的参与在哺乳动物中是保守的。