Department of Physiology, University of Texas Health Science Center, San Antonio, TX 78229, USA.
J Gen Physiol. 2009 Nov;134(5):437-48. doi: 10.1085/jgp.200910313.
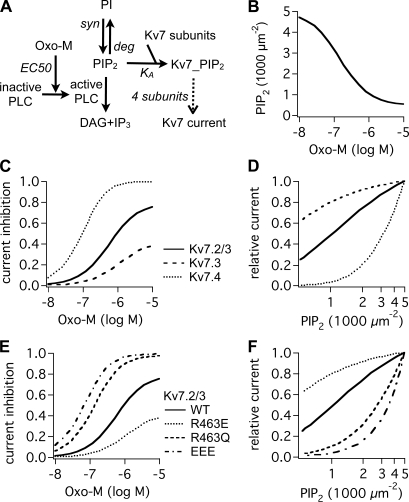
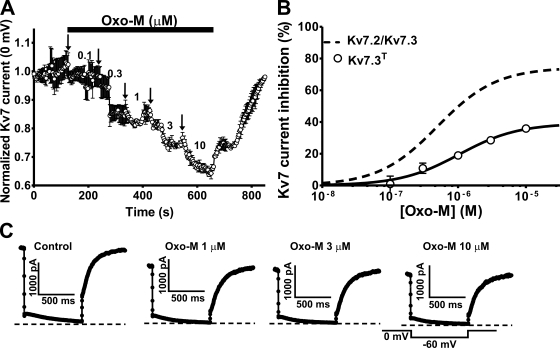
Kv7 K(+)-channel subunits differ in their apparent affinity for PIP(2) and are differentially expressed in nerve, muscle, and epithelia in accord with their physiological roles in those tissues. To investigate how PIP(2) affinity affects the response to physiological stimuli such as receptor stimulation, we exposed homomeric and heteromeric Kv7.2, 7.3, and 7.4 channels to a range of concentrations of the muscarinic receptor agonist oxotremorine-M (oxo-M) in a heterologous expression system. Activation of M(1) receptors by oxo-M leads to PIP(2) depletion through G(q) and phospholipase C (PLC). Chinese hamster ovary cells were transiently transfected with Kv7 subunits and M(1) receptors and studied under perforated-patch voltage clamp. For Kv7.2/7.3 heteromers, the EC(50) for current suppression was 0.44 +/- 0.08 microM, and the maximal inhibition (Inhib(max)) was 74 +/- 3% (n = 5-7). When tonic PIP(2) abundance was increased by overexpression of PIP 5-kinase, the EC(50) was shifted threefold to the right (1.2 +/- 0.1 microM), but without a significant change in Inhib(max) (73 +/- 4%, n = 5). To investigate the muscarinic sensitivity of Kv7.3 homomers, we used the A315T pore mutant (Kv7.3(T)) that increases whole-cell currents by 30-fold without any change in apparent PIP(2) affinity. Kv7.3(T) currents had a slightly right-shifted EC(50) as compared with Kv7.2/7.3 heteromers (1.0 +/- 0.8 microM) and a strongly reduced Inhib(max) (39 +/- 3%). In contrast, the dose-response curve of homomeric Kv7.4 channels was shifted considerably to the left (66 +/- 8 nM), and Inhib(max) was slightly increased (81 +/- 6%, n = 3-4). We then studied several Kv7.2 mutants with altered apparent affinities for PIP(2) by coexpressing them with Kv7.3(T) subunits to boost current amplitudes. For the lower affinity (Kv7.2 (R463Q)/Kv7.3(T)) or higher affinity (Kv7.2 (R463E)/Kv7.3(T)) channels, the EC(50) and Inhib(max) were similar to Kv7.4 or Kv7.3(T) homomers (0.12 +/- 0.08 microM and 79 +/- 6% [n = 3-4] and 0.58 +/- 0.07 microM and 27 +/- 3% [n = 3-4], respectively). The very low-affinity Kv7.2 (R452E, R459E, and R461E) triple mutant was also coexpressed with Kv7.3(T). The resulting heteromer displayed a very low EC(50) for inhibition (32 +/- 8 nM) and a slightly increased Inhib(max) (83 +/- 3%, n = 3-4). We then constructed a cellular model that incorporates PLC activation by oxo-M, PIP(2) hydrolysis, PIP(2) binding to Kv7-channel subunits, and K(+) current through Kv7 tetramers. We were able to fully reproduce our data and extract a consistent set of PIP(2) affinities.
Kv7 K(+) 通道亚基对 PIP(2)的表观亲和力不同,并且根据其在神经、肌肉和上皮组织中的生理作用在这些组织中表现出不同的表达。为了研究 PIP(2)亲和力如何影响对生理刺激(如受体刺激)的反应,我们在异源表达系统中用一系列浓度的毒蕈碱受体激动剂 oxotremorine-M (oxo-M) 暴露同源和异源 Kv7.2、7.3 和 7.4 通道。oxo-M 激活 M(1) 受体通过 G(q)和磷脂酶 C (PLC)导致 PIP(2)耗竭。中国仓鼠卵巢细胞瞬时转染 Kv7 亚基和 M(1)受体,并在穿孔贴片电压钳下进行研究。对于 Kv7.2/7.3 异源二聚体,电流抑制的 EC(50)为 0.44 +/- 0.08 microM,最大抑制 (Inhib(max)) 为 74 +/- 3% (n = 5-7)。当通过过表达 PIP 5-激酶增加 PIP 丰度时,EC(50)向右移动三倍(1.2 +/- 0.1 microM),但 Inhib(max) 没有明显变化(73 +/- 4%,n = 5)。为了研究 Kv7.3 同源二聚体的毒蕈碱敏感性,我们使用了 A315T 孔突变体 (Kv7.3(T)),该突变体使整个细胞电流增加 30 倍,而对表观 PIP(2)亲和力没有任何变化。与 Kv7.2/7.3 异源二聚体相比,Kv7.3(T)电流的 EC(50)略有右移(1.0 +/- 0.8 microM),Inhib(max) 明显降低(39 +/- 3%)。相比之下,同源 Kv7.4 通道的剂量反应曲线向左移动(66 +/- 8 nM),Inhib(max) 略有增加(81 +/- 6%,n = 3-4)。然后,我们通过与 Kv7.3(T) 亚基共表达来研究几种表观 PIP(2)亲和力改变的 Kv7.2 突变体,以提高电流幅度。对于较低亲和力 (Kv7.2 (R463Q)/Kv7.3(T)) 或较高亲和力 (Kv7.2 (R463E)/Kv7.3(T)) 通道,EC(50) 和 Inhib(max) 与 Kv7.4 或 Kv7.3(T) 同源二聚体相似(0.12 +/- 0.08 microM 和 79 +/- 6% [n = 3-4] 和 0.58 +/- 0.07 microM 和 27 +/- 3% [n = 3-4])。非常低亲和力 Kv7.2 (R452E、R459E 和 R461E) 三重突变体也与 Kv7.3(T) 共表达。所得的异源二聚体对抑制具有非常低的 EC(50)(32 +/- 8 nM)和稍高的 Inhib(max)(83 +/- 3%,n = 3-4)。然后,我们构建了一个细胞模型,该模型包含 oxo-M 激活的 PLC、PIP(2)水解、PIP(2)与 Kv7 通道亚基结合以及通过 Kv7 四聚体的 K(+)电流。我们能够完全重现我们的数据并提取一组一致的 PIP(2)亲和力。