Department of Biochemistry and Molecular Biophysics, Washington University Medical School, St Louis, MO 63110, USA.
J Mol Biol. 2010 Mar 19;397(1):260-77. doi: 10.1016/j.jmb.2009.12.051. Epub 2010 Jan 18.
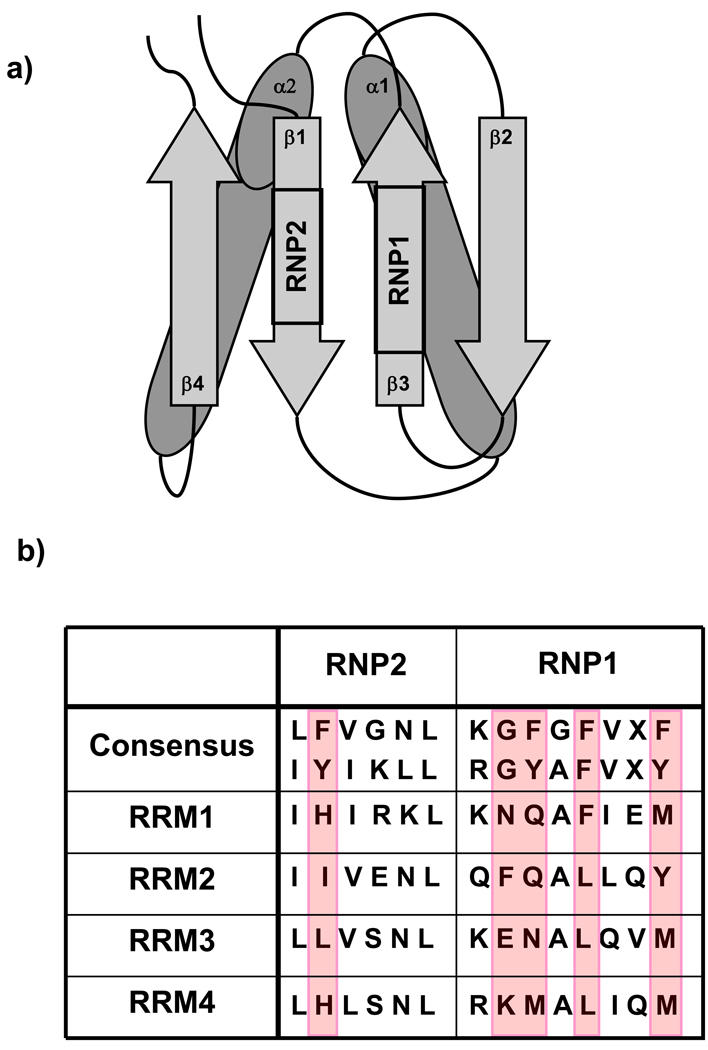
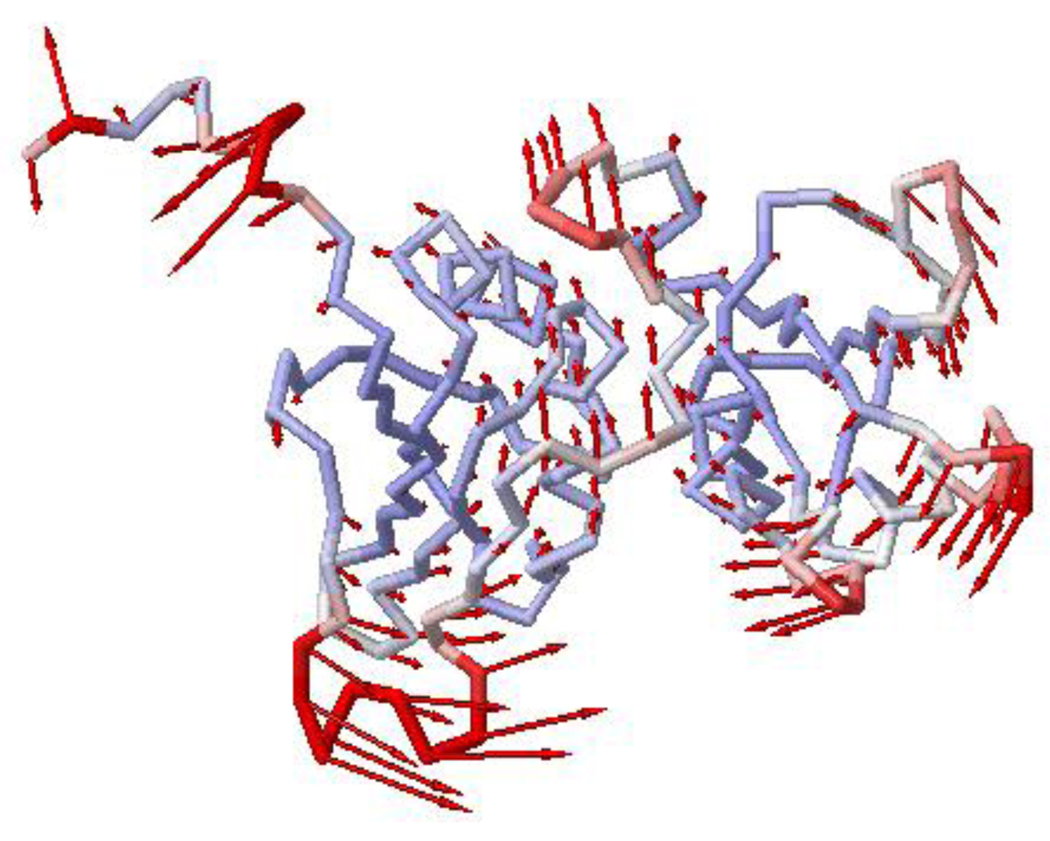
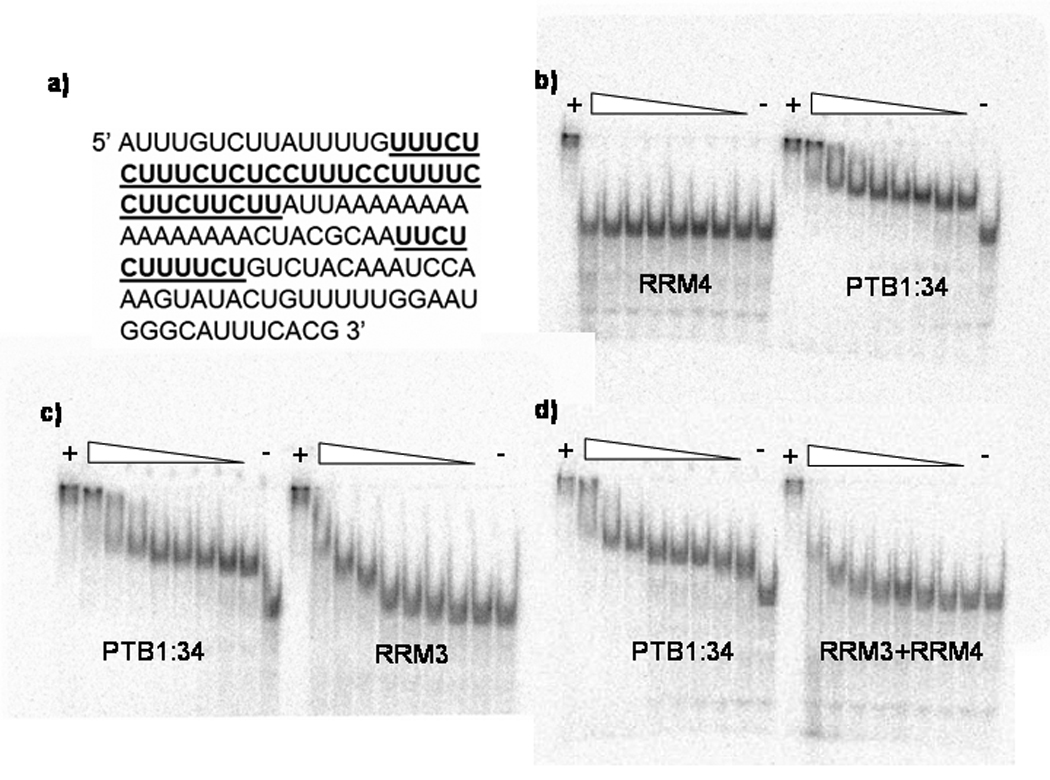
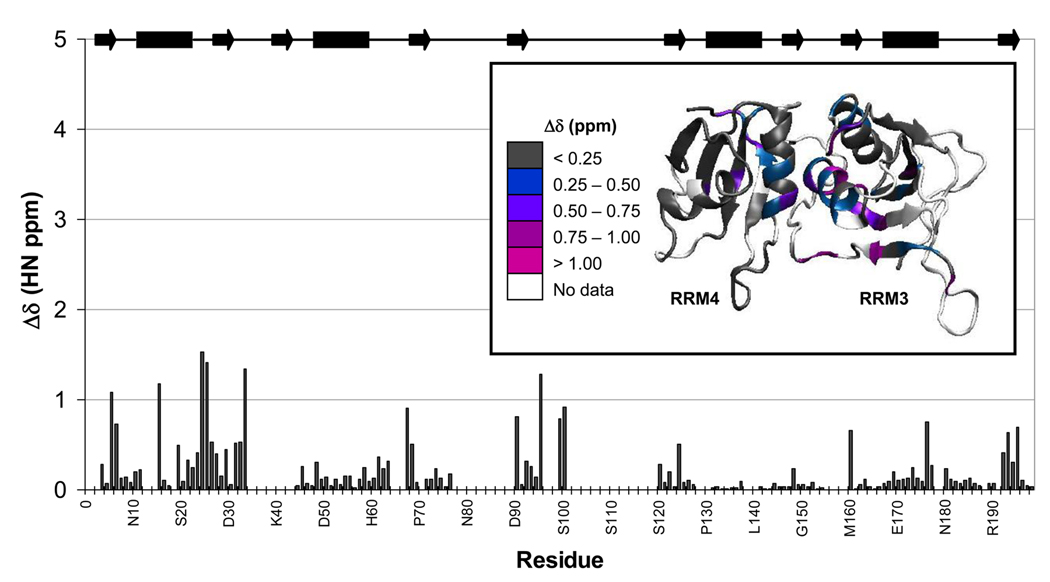
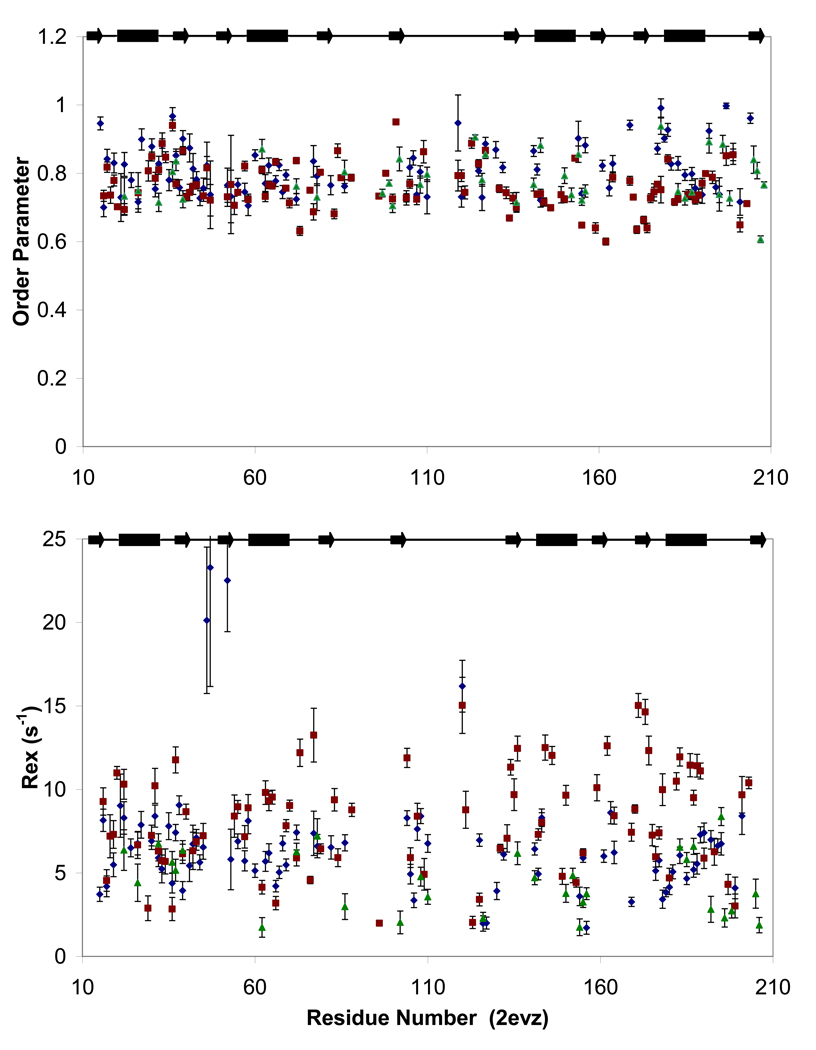
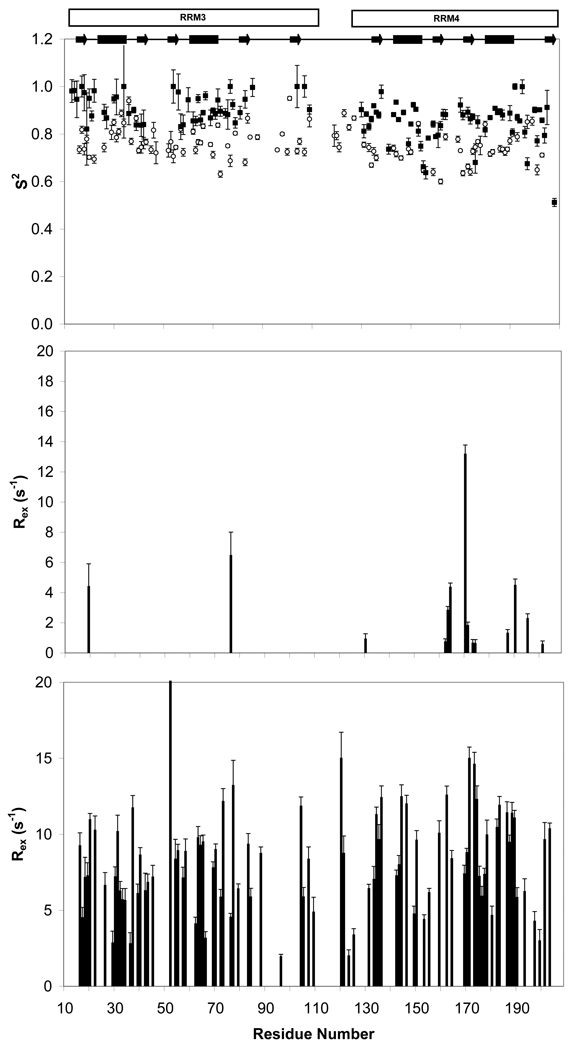
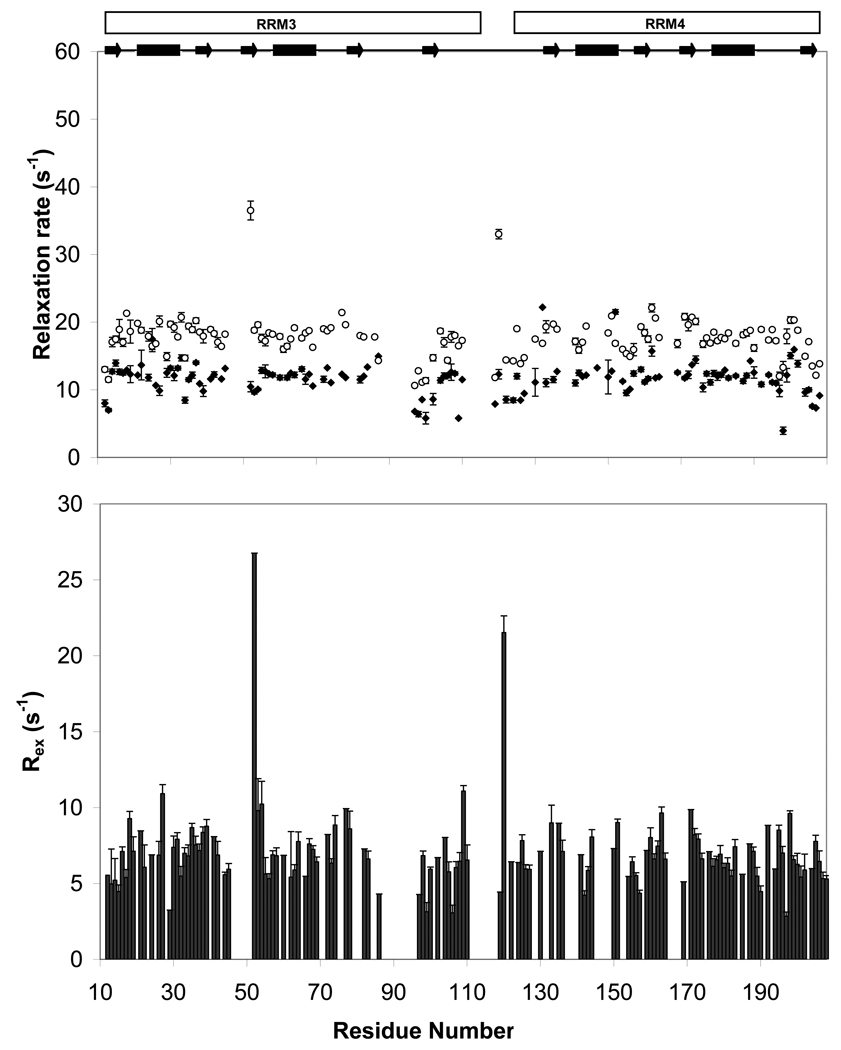
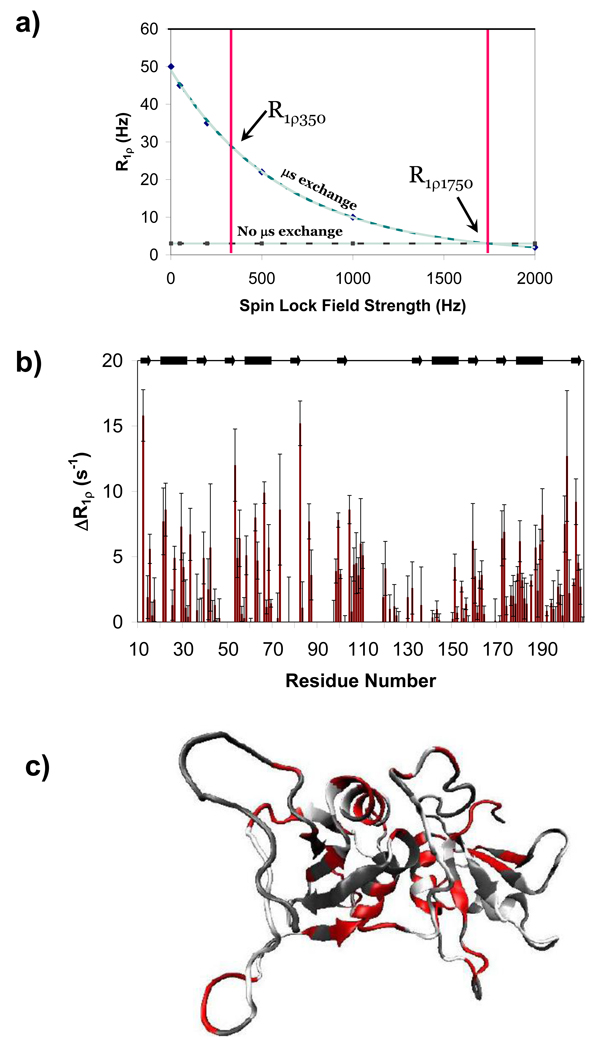
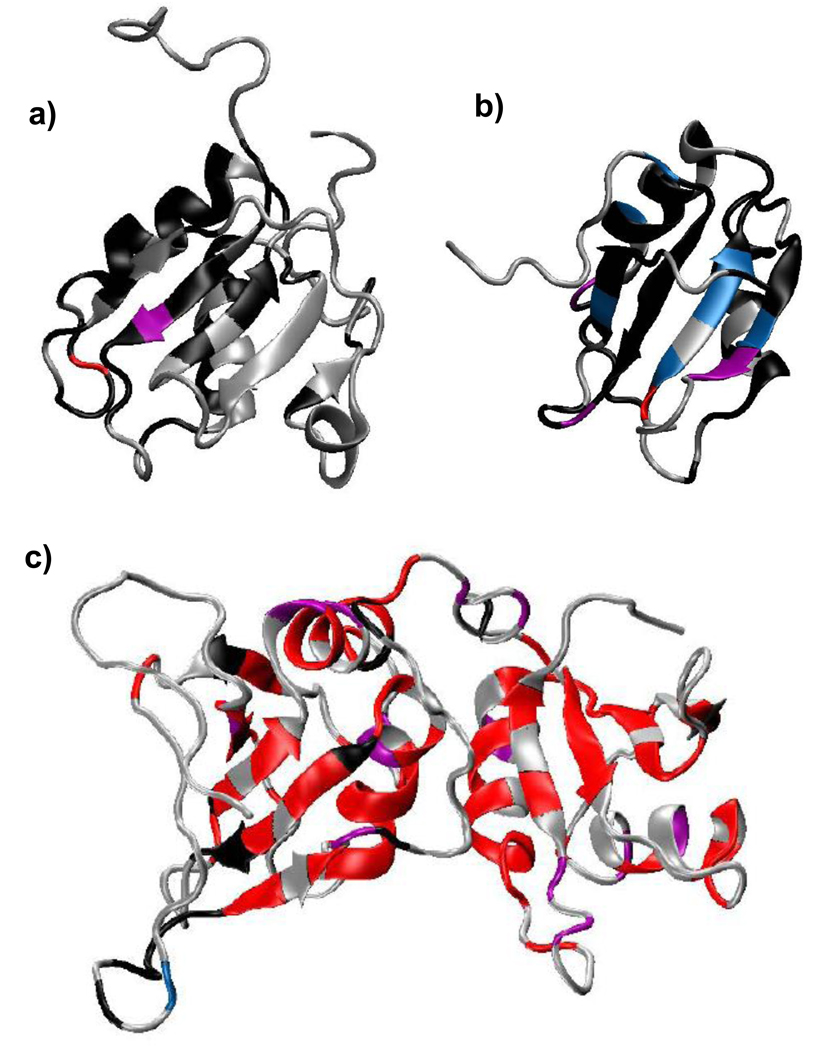
Polypyrimidine tract binding protein (PTB) participates in a variety of functions in eukaryotic cells, including alternative splicing, mRNA stabilization, and internal ribosomal entry site-mediated translation initiation. Its mechanism of RNA recognition is determined in part by the novel geometry of its two C-terminal RNA recognition motifs (RRM3 and RRM4), which interact with each other to form a stable complex (PTB1:34). This complex itself is unusual among RRMs, suggesting that it performs a specific function for the protein. In order to understand the advantage it provides to PTB, the fundamental properties of PTB1:34 are examined here as a comparative study of the complex and its two constituent RRMs. Both RRM3 and RRM4 adopt folded structures that NMR data show to be similar to their structure in PRB1:34. The RNA binding properties of the domains differ dramatically. The affinity of each separate RRM for polypyrimidine tracts is far weaker than that of PTB1:34, and simply mixing the two RRMs does not create an equivalent binding platform. (15)N NMR relaxation experiments show that PTB1:34 has slow, microsecond motions throughout both RRMs including the interdomain linker. This is in contrast to the individual domains, RRM3 and RRM4, where only a few backbone amides are flexible on this time scale. The slow backbone dynamics of PTB1:34, induced by packing of RRM3 and RRM4, could be essential for high-affinity binding to a flexible polypyrimidine tract RNA and also provide entropic compensation for its own formation.
多嘧啶 tract 结合蛋白(PTB)参与真核细胞的多种功能,包括选择性剪接、mRNA 稳定和内部核糖体进入位点介导的翻译起始。其 RNA 识别机制部分取决于其两个 C 端 RNA 识别基序(RRM3 和 RRM4)的新颖几何形状,这两个基序相互作用形成稳定的复合物(PTB1:34)。该复合物本身在 RRMs 中很不常见,表明它为蛋白质执行特定功能。为了了解它为 PTB 提供的优势,本文作为对复合物及其两个组成 RRMs 的比较研究,检查了 PTB1:34 的基本性质。RRM3 和 RRM4 均采用折叠结构,NMR 数据表明其结构类似于 PRB1:34 中的结构。这些结构域的 RNA 结合性质差异很大。每个单独的 RRM 对多嘧啶序列的亲和力远弱于 PTB1:34,并且简单地混合两个 RRMs 并不能创建等效的结合平台。(15)N NMR 弛豫实验表明,PTB1:34 在两个 RRMs 中均具有缓慢的微秒运动,包括结构域间连接。这与单个结构域 RRM3 和 RRM4 形成对比,在这些时间尺度上,只有少数骨架酰胺是灵活的。由 RRM3 和 RRM4 包装诱导的 PTB1:34 的慢骨架动力学可能对于与柔性多嘧啶序列 RNA 的高亲和力结合至关重要,并且为其自身形成提供了熵补偿。