Department of Chemistry and Biochemistry, University of Delaware, Newark, Delaware 19716, USA.
Biochemistry. 2010 Mar 9;49(9):2075-85. doi: 10.1021/bi902222s.
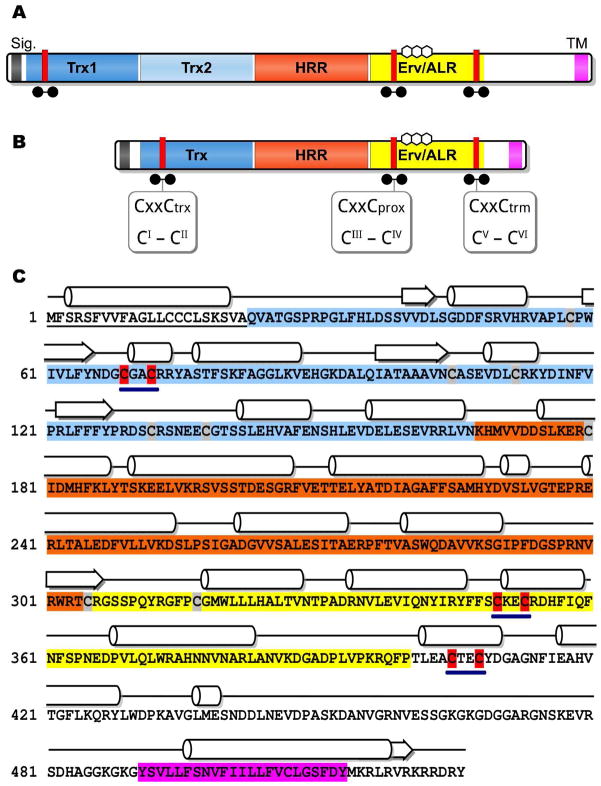
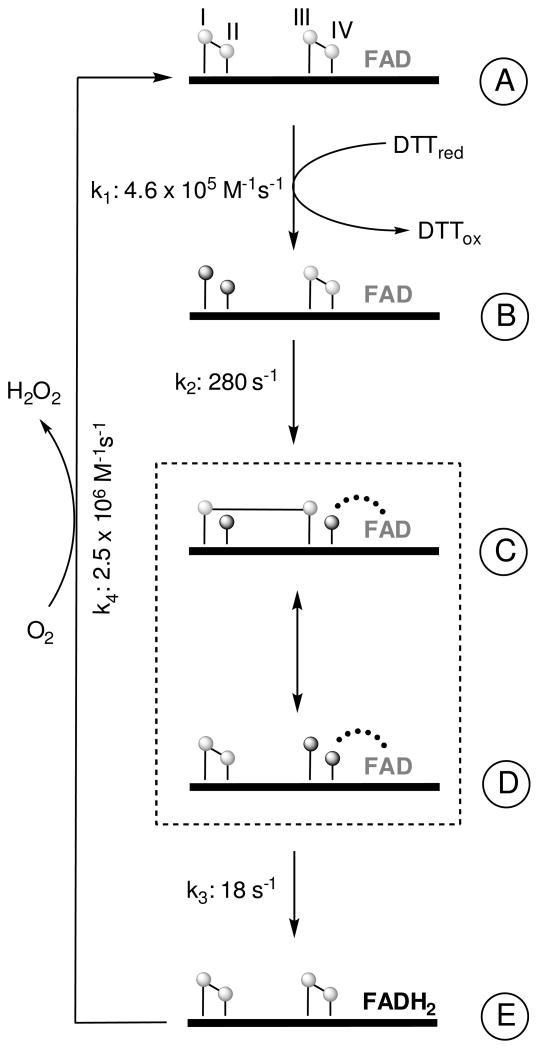
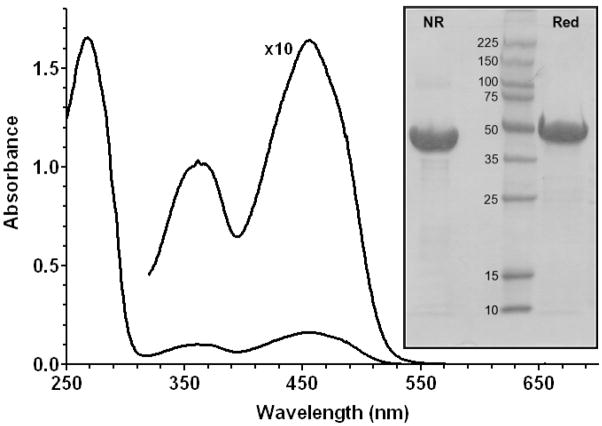
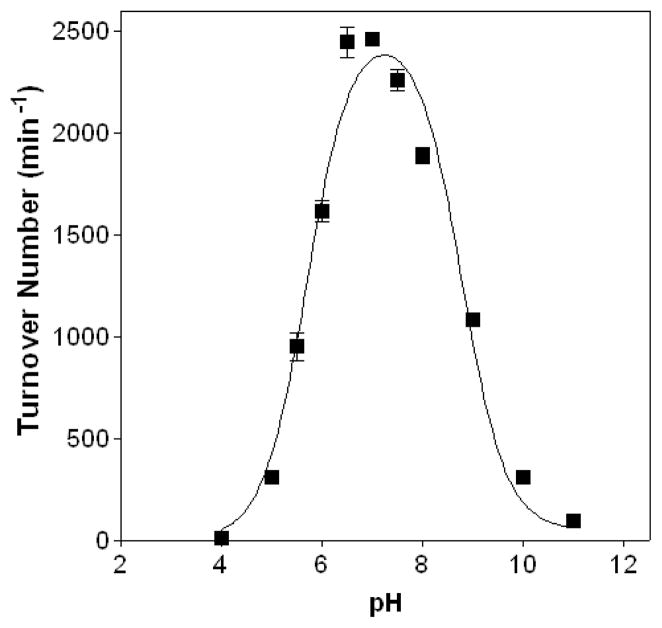
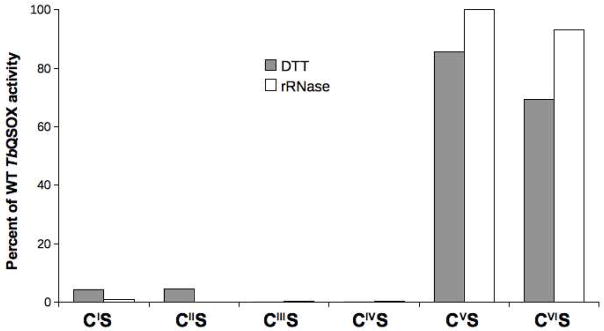
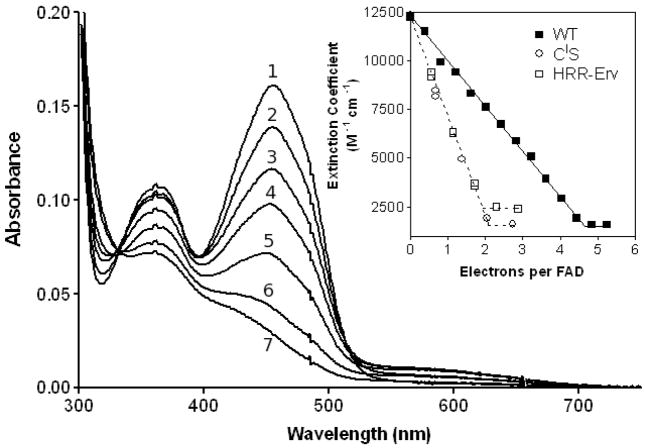
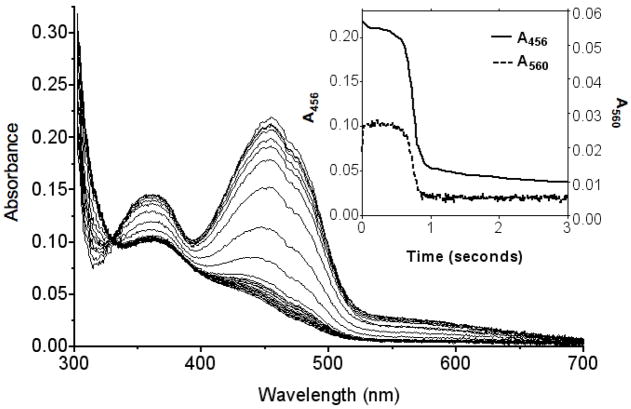
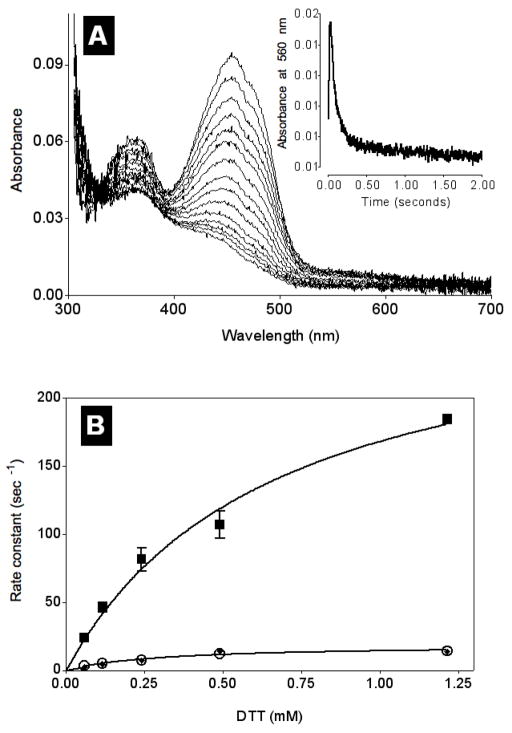
Quiescin sulfhydryl oxidase (QSOX) flavoenzymes catalyze the direct, facile, insertion of disulfide bonds into reduced unfolded proteins with the reduction of oxygen to hydrogen peroxide. To date, only QSOXs from vertebrates have been characterized enzymatically. These metazoan sulfhydryl oxidases have four recognizable domains: a redox-active thioredoxin (Trx) domain containing the first of three CxxC motifs (C(I)-C(II)), a second Trx domain with no obvious redox-active disulfide, a helix-rich domain, and then an Erv/ALR domain. This last domain contains the FAD moiety, a proximal C(III)-C(IV) disulfide, and a third CxxC of unknown function (C(V)-C(VI)). Plant and protist QSOXs lack the second Trx domain but otherwise appear to contain the same complement of redox centers. This work presents the first characterization of a single-Trx QSOX. Trypanosoma brucei QSOX was expressed in Escherichia coli using a synthetic gene and found to be a stable, monomeric, FAD-containing protein. Although evidently lacking an entire domain, TbQSOX shows catalytic activity and substrate specificity similar to the vertebrate QSOXs examined previously. Unfolded reduced proteins are more than 200-fold more effective substrates on a per thiol basis than glutathione and some 10-fold better than the parasite bisglutathione analogue, trypanothione. These data are consistent with a role for the protist QSOX in oxidative protein folding. Site-directed mutagenesis of each of the six cysteine residues (to serines) shows that the CxxC motif in the single-Trx domain is crucial for efficient catalysis of the oxidation of both reduced RNase and the model substrate dithiothreitol. As expected, the proximal disulfide C(III)-C(IV), which interacts with the flavin, is catalytically crucial. However, as observed with human QSOX1, the third CxxC motif shows no obvious catalytic role during the in vitro oxidation of reduced RNase or dithiothreitol. Pre-steady-state kinetics demonstrates that turnover in TbQSOX is limited by an internal redox step leading to 2-electron reduction of the FAD cofactor. In sum, the single-Trx domain QSOX studied here shows a striking similarity in enzymatic behavior to its double-Trx metazoan counterparts.
去氢表雄酮巯基氧化酶 (QSOX) 黄素酶能够催化将二硫键直接、轻松地插入到未折叠的还原态蛋白质中,同时将氧还原为过氧化氢。迄今为止,只有脊椎动物的 QSOX 已被酶法表征。这些后生动物的巯基氧化酶具有四个可识别的结构域:一个包含三个 CxxC 基序(C(I)-C(II))中的第一个的氧化还原活性硫氧还蛋白 (Trx) 结构域,一个没有明显氧化还原活性二硫键的第二个 Trx 结构域,一个富含螺旋的结构域,然后是一个 Erv/ALR 结构域。最后这个结构域包含 FAD 部分、一个近位 C(III)-C(IV) 二硫键和一个功能未知的第三个 CxxC(C(V)-C(VI))。植物和原生动物 QSOX 缺乏第二个 Trx 结构域,但除此之外似乎包含相同的氧化还原中心。这项工作首次对单-Trx QSOX 进行了表征。在大肠杆菌中使用合成基因表达了布氏锥虫 QSOX,发现它是一种稳定的、单体的、含 FAD 的蛋白质。尽管显然缺少整个结构域,但 TbQSOX 表现出与以前研究过的脊椎动物 QSOX 相似的催化活性和底物特异性。在每一个硫醇基础上,未折叠的还原态蛋白质的效力比谷胱甘肽高 200 多倍,比寄生虫双谷胱甘肽类似物 trypanothione 好 10 倍左右。这些数据表明原生动物 QSOX 在氧化蛋白折叠中发挥作用。对六个半胱氨酸残基(突变为丝氨酸)的定点突变表明,单-Trx 结构域中的 CxxC 基序对于有效催化还原 RNase 和模型底物二硫苏糖醇的氧化至关重要。正如预期的那样,与黄素相互作用的近位二硫键 C(III)-C(IV)在催化上是至关重要的。然而,与人类 QSOX1 一样,第三个 CxxC 基序在体外氧化还原 RNase 或二硫苏糖醇时没有明显的催化作用。预稳态动力学表明,TbQSOX 的周转率受到内部氧化还原步骤的限制,导致 FAD 辅因子的 2 电子还原。总之,本文研究的单-Trx 结构域 QSOX 在酶促行为上与其双-Trx 后生动物对应物非常相似。