Department of Biochemistry, University of Oxford, South Parks Road, Oxford OX1 3QU, UK.
Curr Biol. 2010 Feb 23;20(4):279-89. doi: 10.1016/j.cub.2009.12.059. Epub 2010 Feb 11.
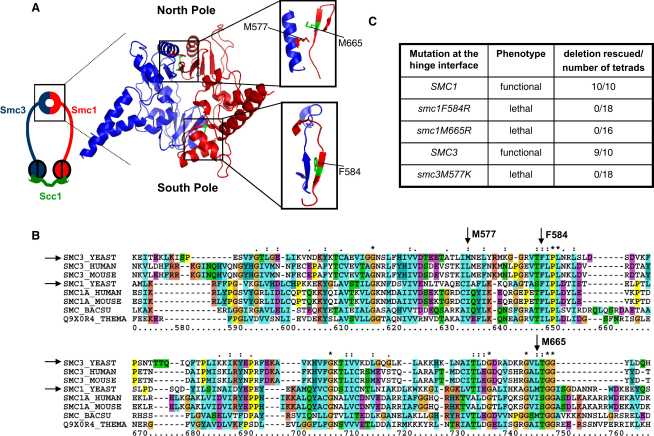
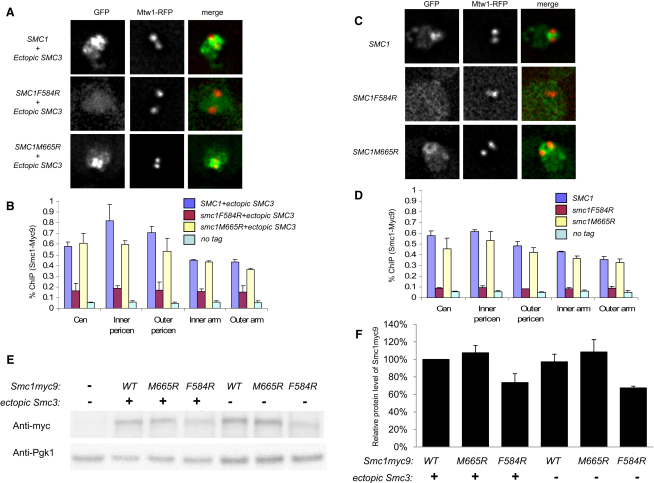
The cohesin complex that mediates sister chromatid cohesion contains three core subunits: Smc1, Smc3, and Scc1. Heterotypic interactions between Smc1 and Smc3 dimerization domains create stable V-shaped Smc1/Smc3 heterodimers with a hinge at the center and nucleotide-binding domains (NBDs) at the ends of each arm. Interconnection of each NBD through their association with the N- and C-terminal domains of Scc1 creates a tripartite ring, within which sister DNAs are thought to be entrapped (the ring model). Crystal structures show that the Smc1/Smc3 hinge has a toroidal shape, with independent "north" and "south" interaction surfaces on an axis of pseudosymmetry. The ring model predicts that sister chromatid cohesion would be lost by transient hinge opening.
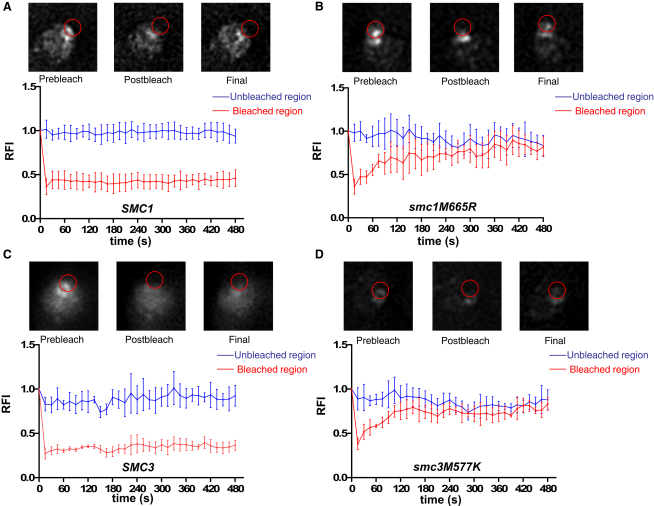
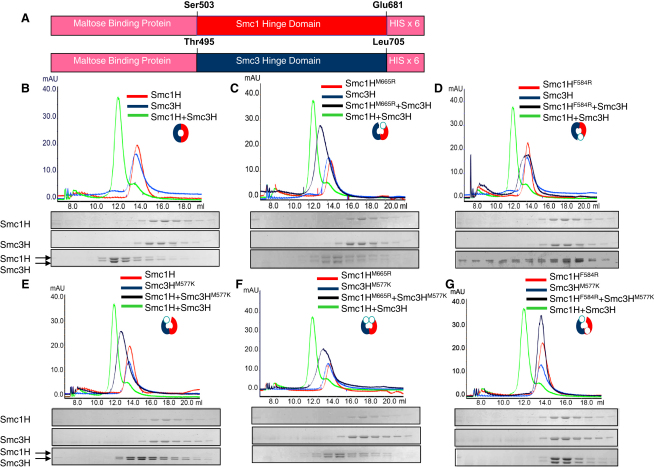
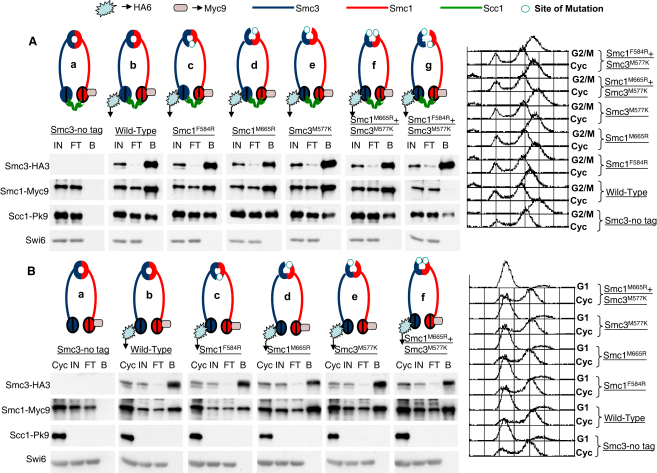
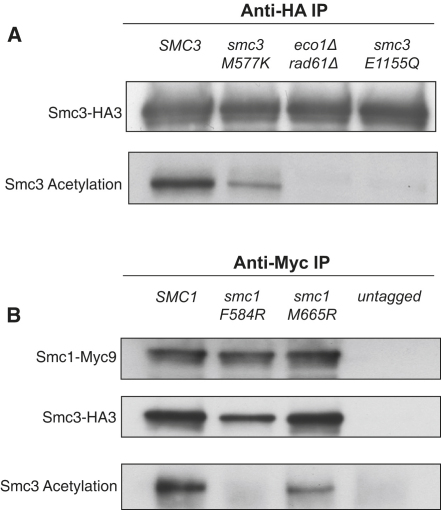
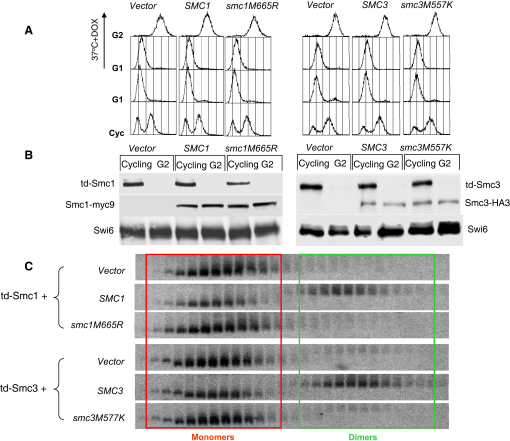
We find that mutations within either interface weaken heterodimerization of isolated half hinges in vitro but do not greatly compromise formation of cohesin rings in vivo. They do, however, reduce the residence time of cohesin on chromosomes and cause lethal defects in sister chromatid cohesion. This demonstrates that mere formation of rings is insufficient for cohesin function. Stable cohesion requires cohesin rings that cannot easily open.
Either the north or south hinge interaction surface is sufficient for the assembly of V-shaped Smc1/Smc3 heterodimers in vivo. Any tendency of Smc proteins with weakened hinges to dissociate will be suppressed by interconnection of their NBDs by Scc1. We suggest that transient hinge dissociation caused by the mutations described here is incompatible with stable sister chromatid cohesion because it permits chromatin fibers to escape from cohesin rings.
介导姐妹染色单体黏合的黏合蛋白复合物包含三个核心亚基:Smc1、Smc3 和 Scc1。Smc1 和 Smc3 二聚化结构域之间的异型相互作用形成稳定的 V 形 Smc1/Smc3 异源二聚体,其中央有一个铰链,每个臂的末端有核苷酸结合结构域(NBD)。每个 NBD 通过与 Scc1 的 N 和 C 末端结构域的连接而相互连接,形成一个三分体环,其中姐妹 DNA 被认为被捕获(环模型)。晶体结构表明,Smc1/Smc3 铰链具有环形形状,在伪对称轴上具有独立的“北”和“南”相互作用表面。环模型预测,姐妹染色单体黏合会因铰链的瞬时打开而丢失。
我们发现,两个界面中的突变都会削弱体外分离的半铰链的异源二聚化,但不会严重影响体内黏合蛋白环的形成。然而,它们会缩短黏合蛋白在染色体上的停留时间,并导致姐妹染色单体黏合的致命缺陷。这表明仅仅形成环不足以发挥黏合蛋白的功能。稳定的黏合需要不易打开的黏合蛋白环。
无论是北方还是南方的铰链相互作用表面,都足以在体内组装 V 形 Smc1/Smc3 异源二聚体。任何带有弱化铰链的 Smc 蛋白的解离倾向都会被 Scc1 连接它们的 NBD 所抑制。我们认为,这里描述的突变所导致的瞬时铰链解离与稳定的姐妹染色单体黏合是不相容的,因为它允许染色质纤维从黏合蛋白环中逃脱。