Department of Physiology, David Geffen School of Medicine, University of California, Los Angeles, Los Angeles, CA 90095, USA.
J Gen Physiol. 2010 May;135(5):415-31. doi: 10.1085/jgp.200910320. Epub 2010 Apr 12.
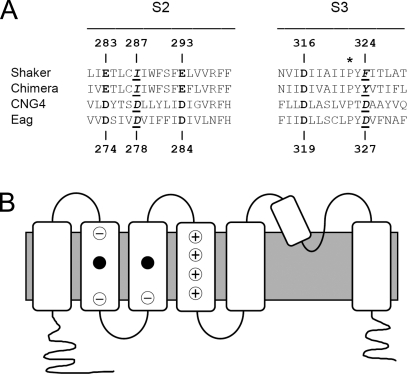
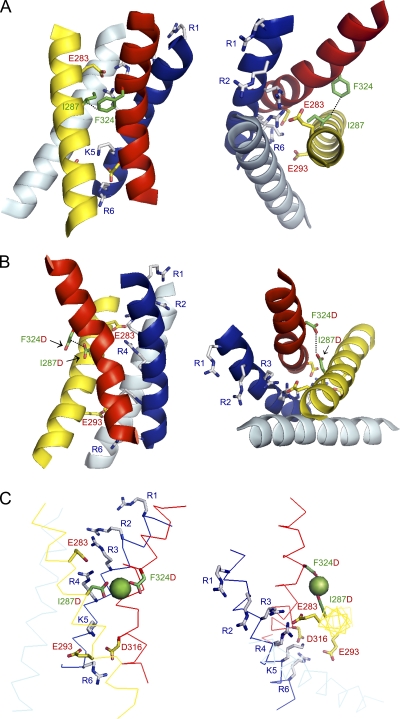
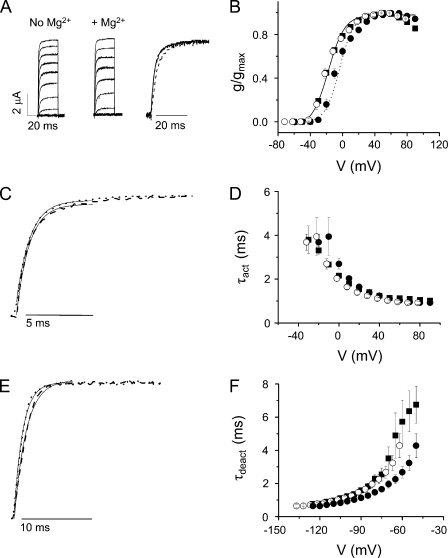
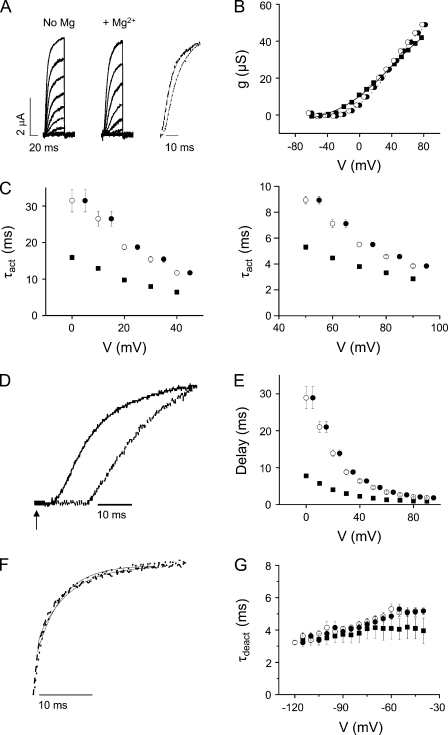
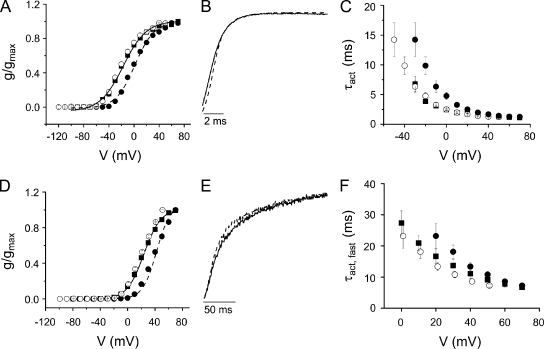
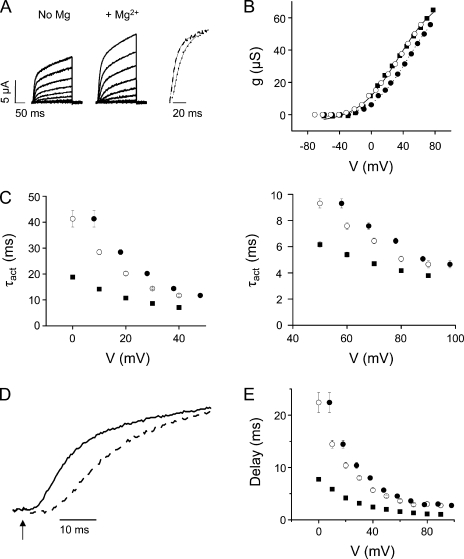
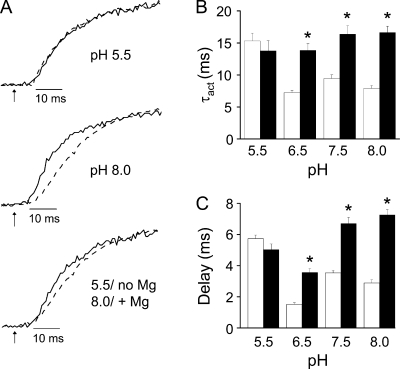
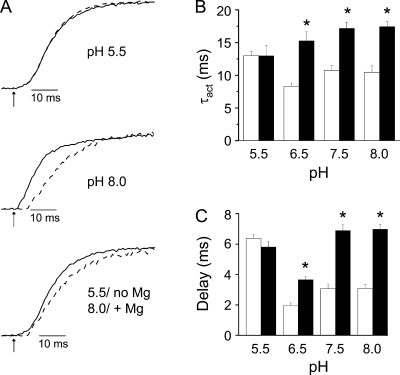
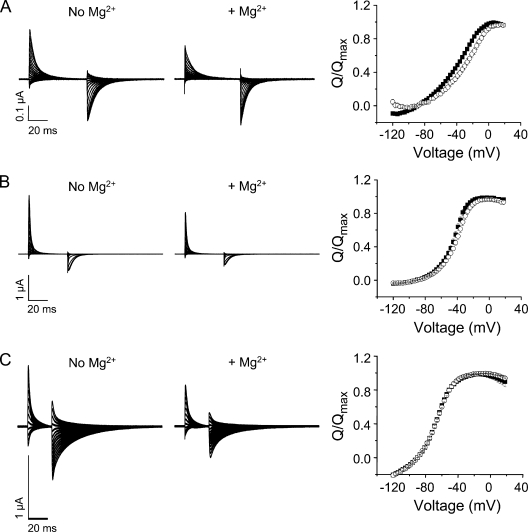
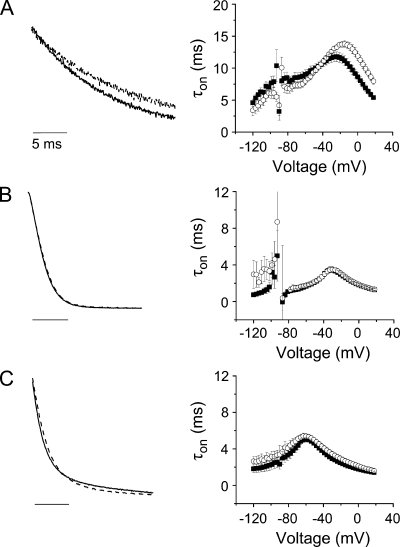
In ether-à-go-go (eag) K(+) channels, extracellular divalent cations bind to the resting voltage sensor and thereby slow activation. Two eag-specific acidic residues in S2 and S3b coordinate the bound ion. Residues located at analogous positions are approximately 4 A apart in the x-ray structure of a Kv1.2/Kv2.1 chimera crystallized in the absence of a membrane potential. It is unknown whether these residues remain in proximity in Kv1 channels at negative voltages when the voltage sensor domain is in its resting conformation. To address this issue, we mutated Shaker residues I287 and F324, which correspond to the binding site residues in eag, to aspartate and recorded ionic and gating currents in the presence and absence of extracellular Mg(2+). In I287D+F324D, Mg(2+) significantly increased the delay before ionic current activation and slowed channel opening with no readily detectable effect on closing. Because the delay before Shaker opening reflects the initial phase of voltage-dependent activation, the results indicate that Mg(2+) binds to the voltage sensor in the resting conformation. Supporting this conclusion, Mg(2+) shifted the voltage dependence and slowed the kinetics of gating charge movement. Both the I287D and F324D mutations were required to modulate channel function. In contrast, E283, a highly conserved residue in S2, was not required for Mg(2+) binding. Ion binding affected activation by shielding the negatively charged side chains of I287D and F324D. These results show that the engineered divalent cation binding site in Shaker strongly resembles the naturally occurring site in eag. Our data provide a novel, short-range structural constraint for the resting conformation of the Shaker voltage sensor and are valuable for evaluating existing models for the resting state and voltage-dependent conformational changes that occur during activation. Comparing our data to the chimera x-ray structure, we conclude that residues in S2 and S3b remain in proximity throughout voltage-dependent activation.
在醚-α- go-go (eag) K(+)通道中,细胞外二价阳离子与静息电压传感器结合,从而减缓激活。S2 和 S3b 中的两个 eag 特异性酸性残基协调结合离子。在不存在膜电位的情况下,用 Kv1.2/Kv2.1 嵌合体的 x 射线结构结晶时,位于 x 射线结构中类似位置的残基之间的距离约为 4A。当电压传感器结构处于静息构象时,Kv1 通道中这些残基在负电压下是否保持接近尚不清楚。为了解决这个问题,我们将 Shaker 残基 I287 和 F324 突变为天冬氨酸,这两个残基对应于 eag 的结合位点残基,并在存在和不存在细胞外 Mg(2+)的情况下记录离子和门控电流。在 I287D+F324D 中,Mg(2+)显著增加了离子电流激活前的延迟,并减缓了通道的开启,对关闭几乎没有可检测的影响。由于 Shaker 开启前的延迟反映了电压依赖性激活的初始阶段,结果表明 Mg(2+)结合在静息构象的电压传感器上。支持这一结论的是,Mg(2+)改变了门控电荷移动的电压依赖性和动力学。I287D 和 F324D 突变都需要调节通道功能。相比之下,S2 中高度保守的残基 E283 不需要与 Mg(2+)结合。离子结合通过屏蔽 I287D 和 F324D 的负电荷侧链来影响激活。这些结果表明,Shaker 中工程化的二价阳离子结合位点强烈类似于 eag 中的天然位点。我们的数据为 Shaker 电压传感器的静息构象提供了一种新颖的短程结构约束,对于评估现有的静息状态和激活过程中发生的电压依赖性构象变化模型非常有价值。将我们的数据与嵌合体 x 射线结构进行比较,我们得出结论,S2 和 S3b 中的残基在整个电压依赖性激活过程中保持接近。