Department of Ecology and Evolutionary Biology, Brown University, Providence, Rhode Island 02912, USA.
Am Nat. 2010 Aug;176(2):227-33. doi: 10.1086/653662.
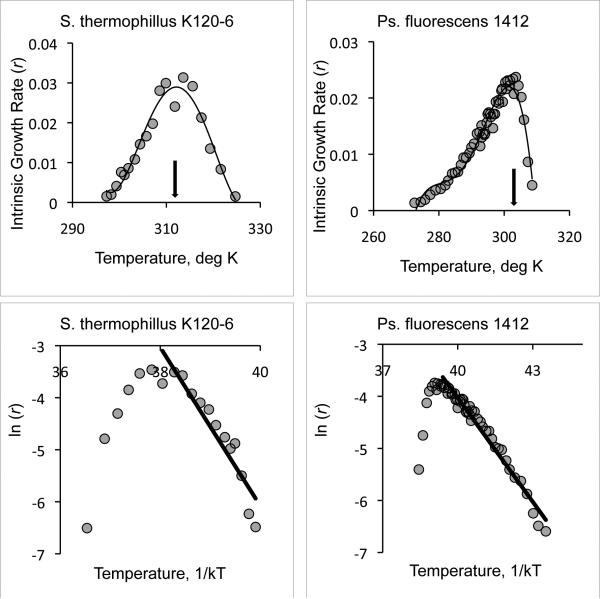
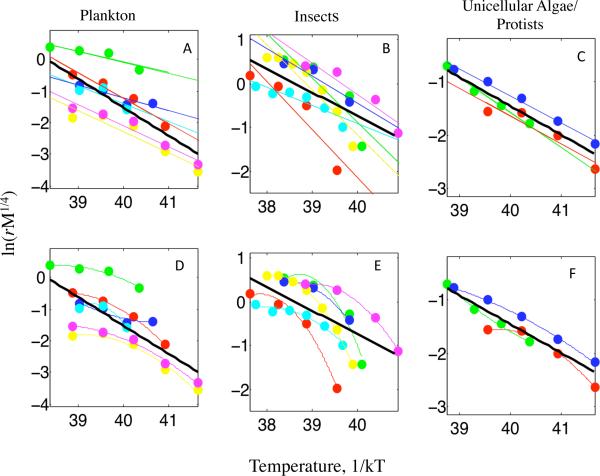
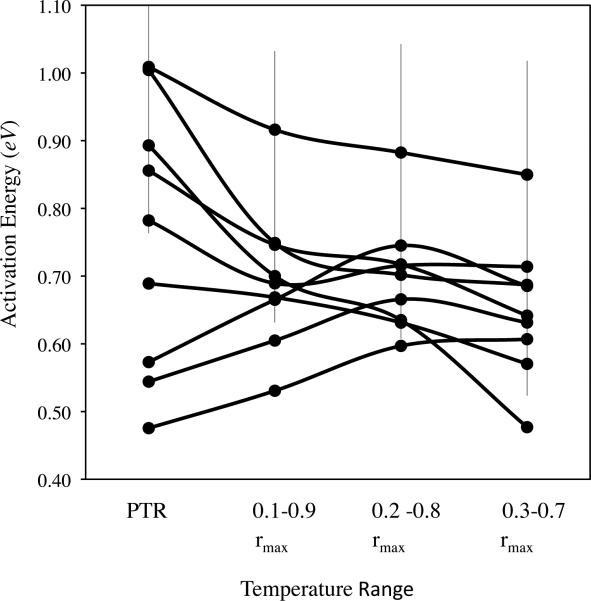
The initial rise of fitness that occurs with increasing temperature is attributed to Arrhenius kinetics, in which rates of reaction increase exponentially with increasing temperature. Models based on Arrhenius typically assume single rate-limiting reactions over some physiological temperature range for which all the rate-limiting enzymes are in 100% active conformation. We test this assumption using data sets for microbes that have measurements of fitness (intrinsic rate of population growth) at many temperatures and over a broad temperature range and for diverse ectotherms that have measurements at fewer temperatures. When measurements are available at many temperatures, strictly Arrhenius kinetics are rejected over the physiological temperature range. However, over a narrower temperature range, we cannot reject strictly Arrhenius kinetics. The temperature range also affects estimates of the temperature dependence of fitness. These results indicate that Arrhenius kinetics only apply over a narrow range of temperatures for ectotherms, complicating attempts to identify general patterns of temperature dependence.
随着温度的升高,最初的健身效果归因于阿仑尼乌斯动力学,其中反应速率随温度呈指数增长。基于阿仑尼乌斯的模型通常假设在某些生理温度范围内存在单一的限速反应,对于所有限速酶,其 100%处于活性构象。我们使用来自微生物的数据来检验这一假设,这些微生物在许多温度下和广泛的温度范围内测量了适应性(种群增长的内在速率),并且对于具有较少温度测量的多种变温动物也是如此。当在许多温度下进行测量时,在生理温度范围内严格的阿仑尼乌斯动力学被拒绝。然而,在更窄的温度范围内,我们不能拒绝严格的阿仑尼乌斯动力学。温度范围还会影响对适应性对温度的依赖性的估计。这些结果表明,阿仑尼乌斯动力学仅适用于变温动物的狭窄温度范围,这使得确定温度依赖性的一般模式变得复杂。