Istituto di Biofisica, Consiglio Nazionale delle Ricerche, 16149 Genoa, Italy.
J Gen Physiol. 2010 Sep;136(3):311-23. doi: 10.1085/jgp.201010455.
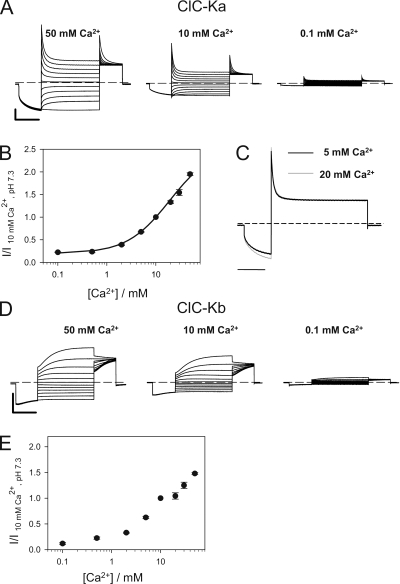
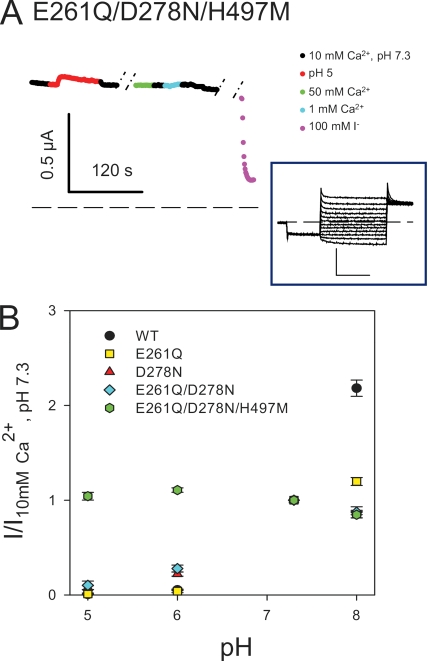
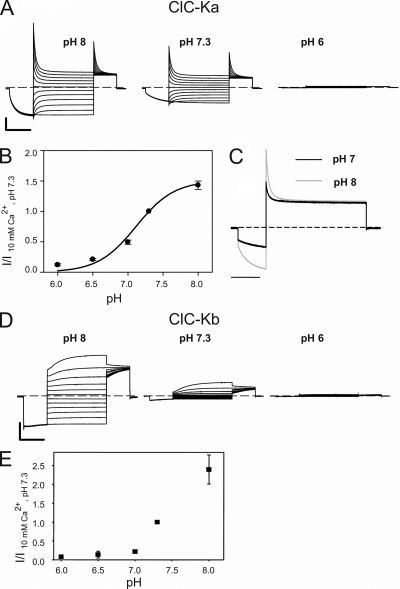
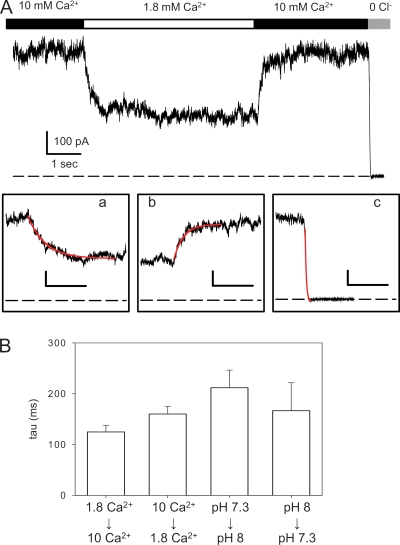
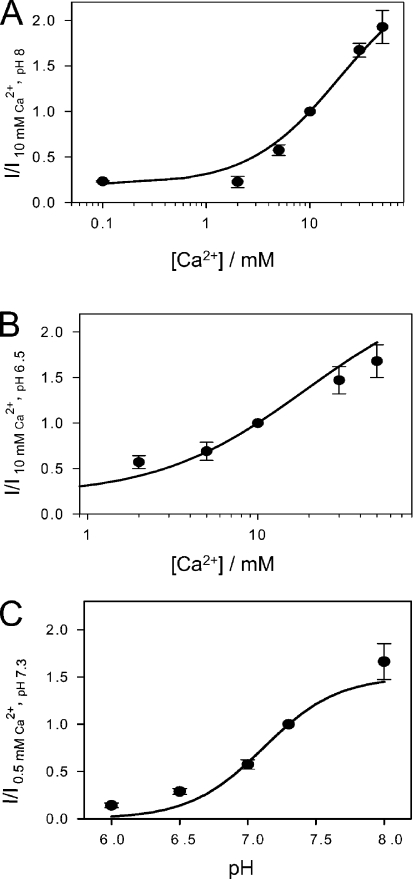
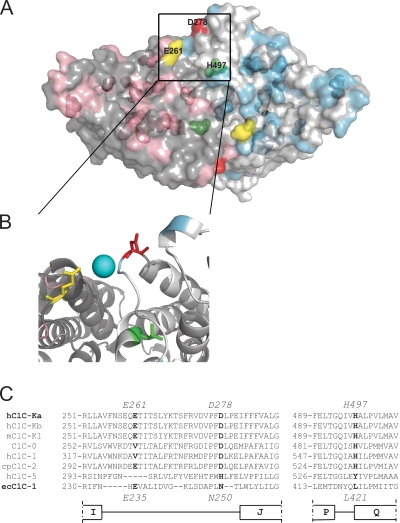
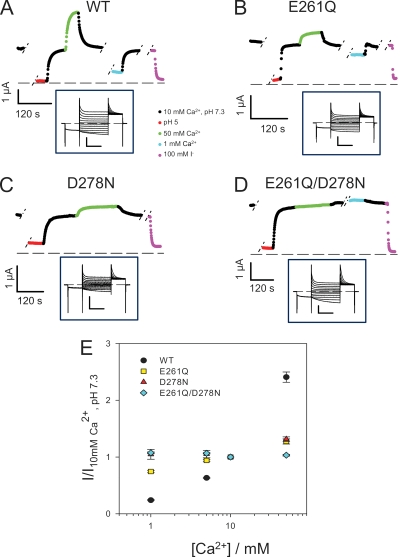
The two human CLC Cl(-) channels, ClC-Ka and ClC-Kb, are almost exclusively expressed in kidney and inner ear epithelia. Mutations in the genes coding for ClC-Kb and barttin, an essential CLC-K channel beta subunit, lead to Bartter syndrome. We performed a biophysical analysis of the modulatory effect of extracellular Ca(2+) and H(+) on ClC-Ka and ClC-Kb in Xenopus oocytes. Currents increased with increasing Ca(2+) without full saturation up to 50 mM. However, in the absence of Ca(2+), ClC-Ka currents were still 20% of currents in 10 mM Ca(2+), demonstrating that Ca(2+) is not strictly essential for opening. Vice versa, ClC-Ka and ClC-Kb were blocked by increasing H(+) with a practically complete block at pH 6. Ca(2+) and H(+) act as gating modifiers without changing the single-channel conductance. Dose-response analysis suggested that two protons are necessary to induce block with an apparent pK of approximately 7.1. A simple four-state allosteric model described the modulation by Ca(2+) assuming a 13-fold higher Ca(2+) affinity of the open state compared with the closed state. The quantitative analysis suggested separate binding sites for Ca(2+) and H(+). A mutagenic screen of a large number of extracellularly accessible amino acids identified a pair of acidic residues (E261 and D278 on the loop connecting helices I and J), which are close to each other but positioned on different subunits of the channel, as a likely candidate for forming an intersubunit Ca(2+)-binding site. Single mutants E261Q and D278N greatly diminished and the double mutant E261Q/D278N completely abolished modulation by Ca(2+). Several mutations of a histidine residue (H497) that is homologous to a histidine that is responsible for H(+) block in ClC-2 did not yield functional channels. However, the triple mutant E261Q/D278N/H497M completely eliminated H(+) -induced current block. We have thus identified a protein region that is involved in binding these physiologically important ligands and that is likely undergoing conformational changes underlying the complex gating of CLC-K channels.
两种人类 CLC Cl(-) 通道,CLC-Ka 和 CLC-Kb,几乎仅在肾脏和内耳上皮细胞中表达。编码 CLC-Kb 和 barttin 的基因突变, barttin 是 CLC-K 通道的必需β亚基,导致 Bartter 综合征。我们在非洲爪蟾卵母细胞中对细胞外 Ca(2+) 和 H(+) 对 ClC-Ka 和 ClC-Kb 的调节作用进行了生物物理分析。随着 Ca(2+) 的增加,电流增加,直至达到 50 mM 而没有完全饱和。然而,在没有 Ca(2+) 的情况下,ClC-Ka 电流仍然是 10 mM Ca(2+) 中电流的 20%,表明 Ca(2+) 对于打开不是严格必需的。反之,ClC-Ka 和 ClC-Kb 被增加的 H(+) 阻断,在 pH 6 时几乎完全阻断。Ca(2+) 和 H(+) 作为门控修饰剂起作用,而不改变单通道电导。剂量反应分析表明,两个质子需要诱导阻断,表观 pK 约为 7.1。一个简单的四态变构模型描述了 Ca(2+) 的调节,假设与关闭状态相比,开放状态的 Ca(2+) 亲和力高 13 倍。定量分析表明,Ca(2+) 和 H(+) 具有单独的结合位点。对大量细胞外可及的氨基酸进行的诱变筛选确定了一对酸性残基(连接 I 和 J 螺旋的环上的 E261 和 D278),它们彼此靠近,但位于通道的不同亚基上,作为形成亚基间 Ca(2+) 结合位点的可能候选物。单个突变 E261Q 和 D278N 大大减少,而双突变 E261Q/D278N 完全消除了 Ca(2+) 的调节作用。一个组氨酸残基(H497)的几个突变,该残基与负责 ClC-2 中 H(+) 阻断的组氨酸同源,没有产生功能性通道。然而,三重突变 E261Q/D278N/H497M 完全消除了 H(+) 诱导的电流阻断。因此,我们确定了一个参与结合这些生理上重要配体的蛋白质区域,并且该区域可能正在经历构成 CLC-K 通道复杂门控的构象变化。