Groningen Biomolecular Sciences and Biotechnology Institute, University of Groningen, Nijenborgh 4, 9747 AG Groningen, The Netherlands.
J Am Chem Soc. 2010 Dec 1;132(47):17004-14. doi: 10.1021/ja107410x. Epub 2010 Nov 8.
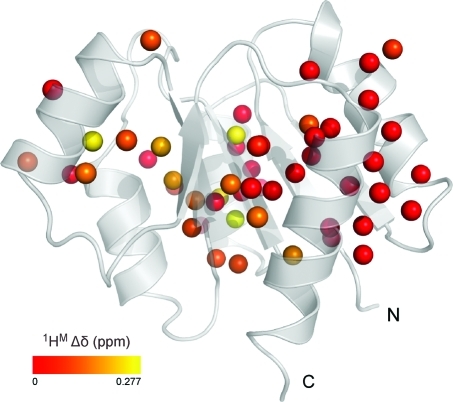
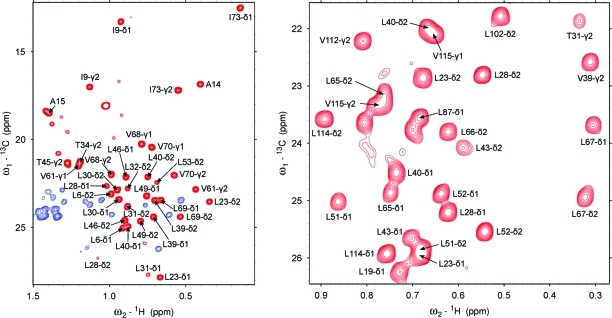
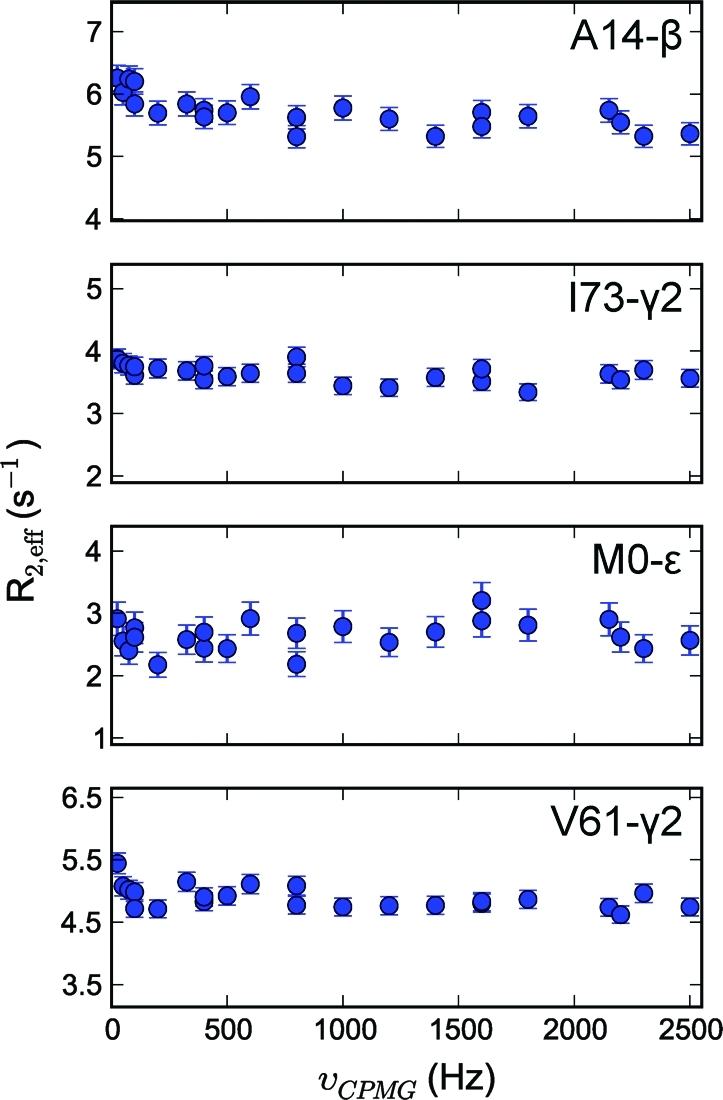
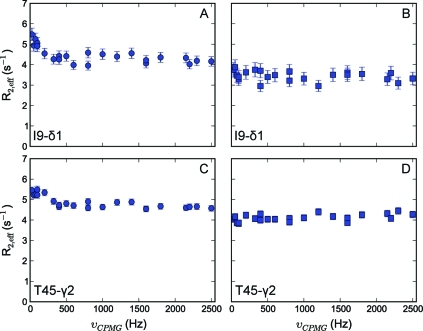
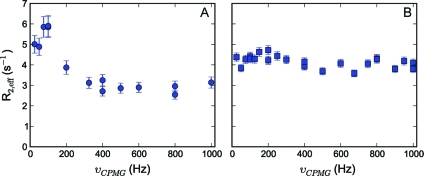
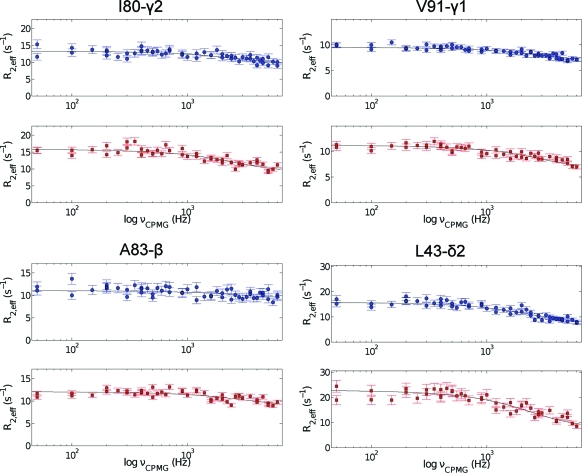
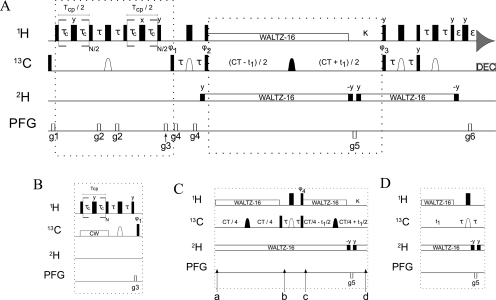
To study microsecond processes by relaxation dispersion NMR spectroscopy, low power deposition and short pulses are crucial and encourage the development of experiments that employ (1)H Carr-Purcell-Meiboom-Gill (CPMG) pulse trains. Herein, a method is described for the comprehensive study of microsecond to millisecond time scale dynamics of methyl groups in proteins, exploiting their high abundance and favorable relaxation properties. In our approach, protein samples are produced using [(1)H, (13)C]-d-glucose in ∼100% D(2)O, which yields CHD(2) methyl groups for alanine, valine, threonine, isoleucine, leucine, and methionine residues with high abundance, in an otherwise largely deuterated background. Methyl groups in such samples can be sequence-specifically assigned to near completion, using (13)C TOCSY NMR spectroscopy, as was recently demonstrated (Otten, R.; et al. J. Am. Chem. Soc. 2010, 132, 2952-2960). In this Article, NMR pulse schemes are presented to measure (1)H CPMG relaxation dispersion profiles for CHD(2) methyl groups, in a vein similar to that of backbone relaxation experiments. Because of the high deuteration level of methyl-bearing side chains, artifacts arising from proton scalar coupling during the CPMG pulse train are negligible, with the exception of Ile-δ1 and Thr-γ2 methyl groups, and a pulse scheme is described to remove the artifacts for those residues. Strong (13)C scalar coupling effects, observed for several leucine residues, are removed by alternative biochemical and NMR approaches. The methodology is applied to the transcriptional activator NtrC(r), for which an inactive/active state transition was previously measured and the motions in the microsecond time range were estimated through a combination of backbone (15)N CPMG dispersion NMR spectroscopy and a collection of experiments to determine the exchange-free component to the transverse relaxation rate. Exchange contributions to the (1)H line width were detected for 21 methyl groups, and these probes were found to collectively report on a local structural rearrangement around the phosphorylation site, with a rate constant of (15.5 ± 0.5) × 10(3) per second (i.e., τ(ex) = 64.7 ± 1.9 μs). The affected methyl groups indicate that, already before phosphorylation, a substantial, transient rearrangement takes place between helices 3 and 4 and strands 4 and 5. This conformational equilibrium allows the protein to gain access to the active, signaling state in the absence of covalent modification through a shift in a pre-existing dynamic equilibrium. Moreover, the conformational switching maps exactly to the regions that differ between the solution NMR structures of the fully inactive and active states. These results demonstrate that a cost-effective and quantitative study of protein methyl group dynamics by (1)H CPMG relaxation dispersion NMR spectroscopy is possible and can be applied to study functional motions on the microsecond time scale that cannot be accessed by backbone (15)N relaxation dispersion NMR. The use of methyl groups as dynamics probes extends such applications also to larger proteins.
为了通过弛豫色散 NMR 光谱研究微秒过程,低功率沉积和短脉冲至关重要,这鼓励了采用(1)H Carr-Purcell-Meiboom-Gill(CPMG)脉冲串的实验的发展。在此,描述了一种用于研究蛋白质中甲基基团在微秒到毫秒时间尺度上的动力学的综合方法,利用了它们的高丰度和良好的弛豫特性。在我们的方法中,使用(1)H,(13)C]-d-葡萄糖在约 100%D(2)O 中生产蛋白质样品,对于丙氨酸、缬氨酸、苏氨酸、异亮氨酸、亮氨酸和甲硫氨酸残基,生成具有高丰度的 CHD(2)甲基基团,在其他情况下,背景主要是氘代的。最近已经证明(Otten,R.等人,J. Am. Chem. Soc. 2010,132,2952-2960),可以使用(13)C TOCSY NMR 光谱对这种样品中的甲基基团进行序列特异性分配,接近完全分配。在本文中,提出了 NMR 脉冲方案,以测量 CHD(2)甲基基团的(1)H CPMG 弛豫色散曲线,其方式类似于 backbone relaxation 实验。由于侧链甲基具有高氘化水平,除了 Ile-δ1 和 Thr-γ2 甲基基团外,在 CPMG 脉冲串期间由质子标量耦合引起的伪影可以忽略不计,并且描述了一种用于去除这些残基伪影的脉冲方案。几个亮氨酸残基中观察到的强(13)C 标量耦合效应通过替代生化和 NMR 方法去除。该方法应用于转录激活剂 NtrC(r),先前已经测量了其无活性/活性状态的转变,并且通过组合 backbone(15)N CPMG 弥散 NMR 光谱和一组实验来确定横向弛豫率的交换自由分量来估计微秒时间范围内的运动。已经检测到 21 个甲基基团的(1)H 线宽的交换贡献,并且这些探针被发现共同报告了围绕磷酸化位点的局部结构重排,其速率常数为(15.5±0.5)×10(3)每秒(即 τ(ex)= 64.7±1.9 μs)。受影响的甲基基团表明,在磷酸化之前,已经在螺旋 3 和 4 以及链 4 和 5 之间发生了大量的瞬时重排。这种构象平衡允许蛋白质在没有共价修饰的情况下通过预先存在的动态平衡的移动获得活性信号状态。此外,构象转换完全映射到完全无活性和活性状态之间溶液 NMR 结构差异的区域。这些结果表明,通过(1)H CPMG 弛豫色散 NMR 光谱对蛋白质甲基基团动力学进行具有成本效益和定量的研究是可能的,并且可以应用于研究无法通过 backbone(15)N 弛豫色散 NMR 访问的微秒时间尺度上的功能运动。将甲基基团用作动力学探针还将这些应用扩展到更大的蛋白质。