Institut de Recherches Cliniques de Montréal, Montréal, Québec, Canada.
PLoS Genet. 2010 Oct 28;6(10):e1001173. doi: 10.1371/journal.pgen.1001173.
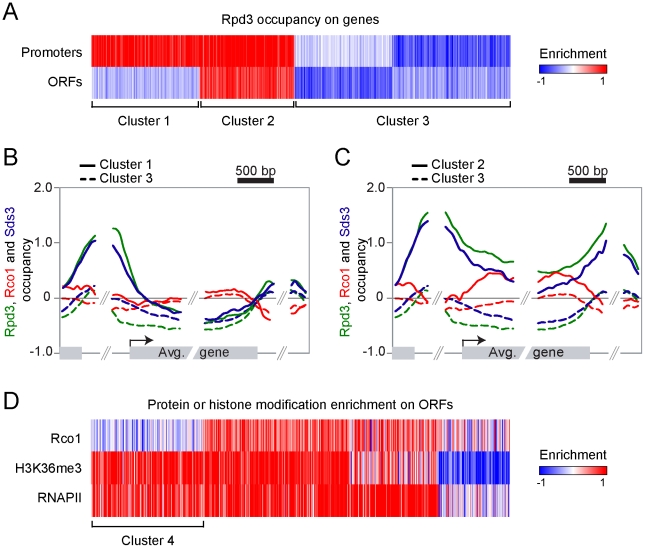
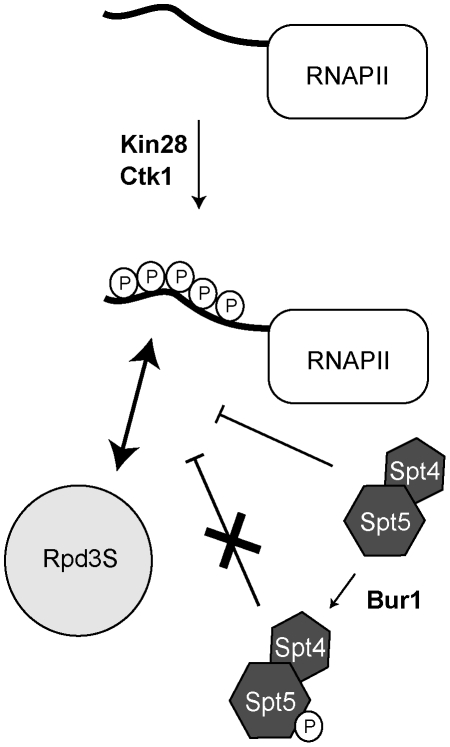
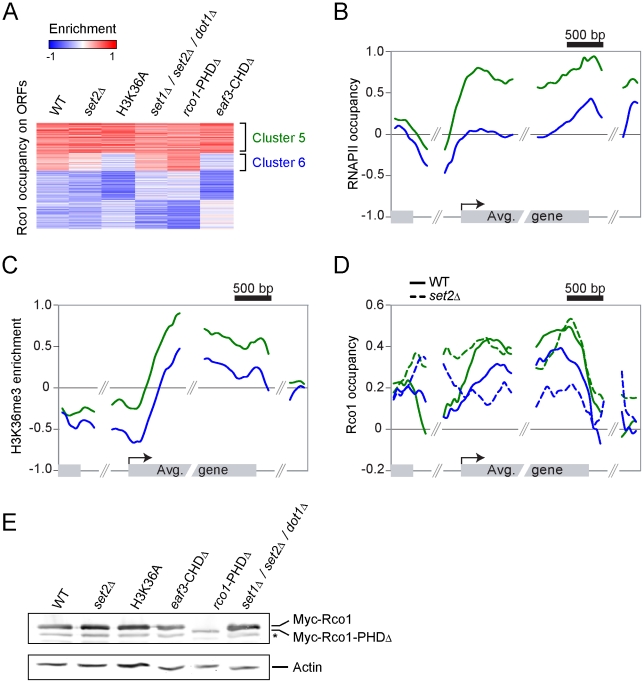
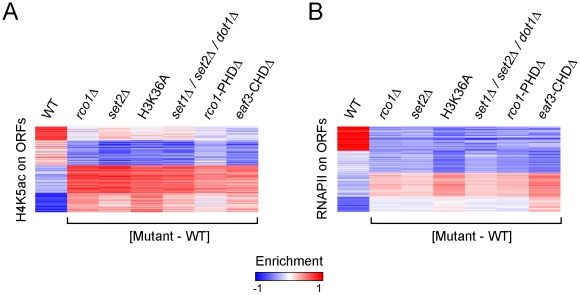
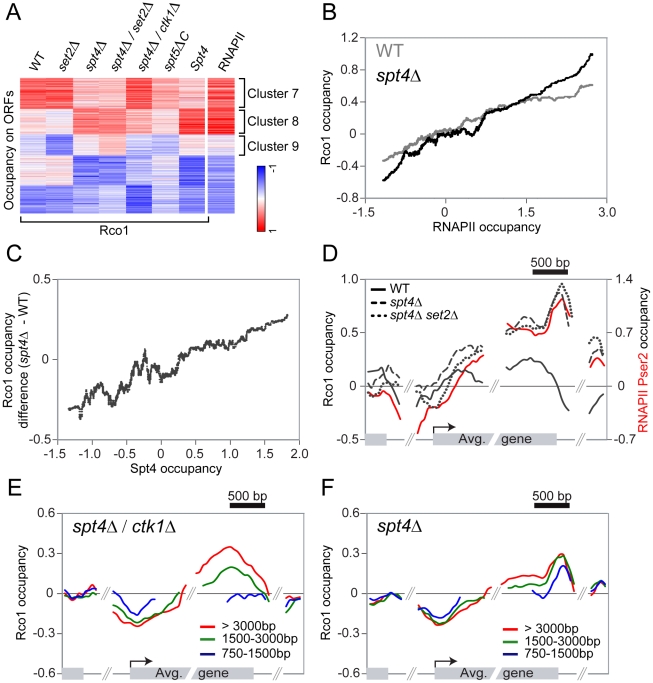
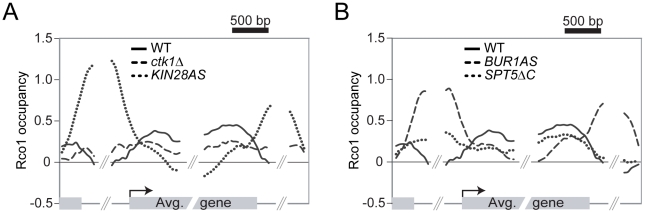
Histone deacetylase Rpd3 is part of two distinct complexes: the large (Rpd3L) and small (Rpd3S) complexes. While Rpd3L targets specific promoters for gene repression, Rpd3S is recruited to ORFs to deacetylate histones in the wake of RNA polymerase II, to prevent cryptic initiation within genes. Methylation of histone H3 at lysine 36 by the Set2 methyltransferase is thought to mediate the recruitment of Rpd3S. Here, we confirm by ChIP-Chip that Rpd3S binds active ORFs. Surprisingly, however, Rpd3S is not recruited to all active genes, and its recruitment is Set2-independent. However, Rpd3S complexes recruited in the absence of H3K36 methylation appear to be inactive. Finally, we present evidence implicating the yeast DSIF complex (Spt4/5) and RNA polymerase II phosphorylation by Kin28 and Ctk1 in the recruitment of Rpd3S to active genes. Taken together, our data support a model where Set2-dependent histone H3 methylation is required for the activation of Rpd3S following its recruitment to the RNA polymerase II C-terminal domain.
组蛋白去乙酰化酶 Rpd3 是两个不同复合物的一部分:大(Rpd3L)和小(Rpd3S)复合物。虽然 Rpd3L 针对特定的启动子进行基因抑制,但 Rpd3S 被招募到 ORF 以在 RNA 聚合酶 II 之后去乙酰化组蛋白,以防止基因内的隐匿起始。组蛋白 H3 赖氨酸 36 的甲基化被认为介导了 Rpd3S 的募集。在这里,我们通过 ChIP-Chip 证实了 Rpd3S 结合活性 ORF。然而,令人惊讶的是,Rpd3S 并非被招募到所有活性基因,并且其募集与 Set2 无关。然而,在没有 H3K36 甲基化的情况下募集的 Rpd3S 复合物似乎没有活性。最后,我们提出了证据表明酵母 DSIF 复合物(Spt4/5)和 RNA 聚合酶 II 由 Kin28 和 Ctk1 磷酸化参与 Rpd3S 向活性基因的募集。总之,我们的数据支持了这样的模型,即依赖于 Set2 的组蛋白 H3 甲基化是 Rpd3S 在被募集到 RNA 聚合酶 II C 末端结构域之后激活所必需的。