Department of Cancer Biology and Irell and Manella Graduate School of Biological Sciences, Beckman Research Institute of the City of Hope, Duarte, California, United States of America.
PLoS Genet. 2010 Nov 4;6(11):e1001194. doi: 10.1371/journal.pgen.1001194.
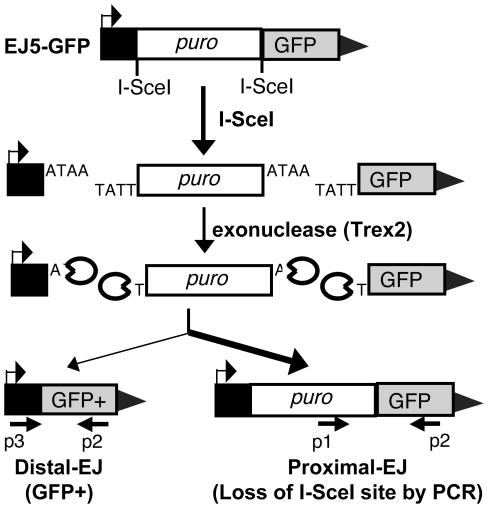
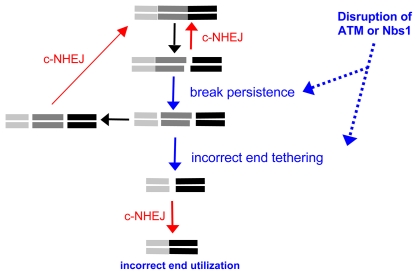
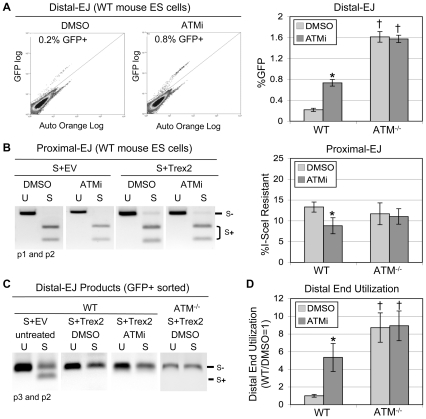
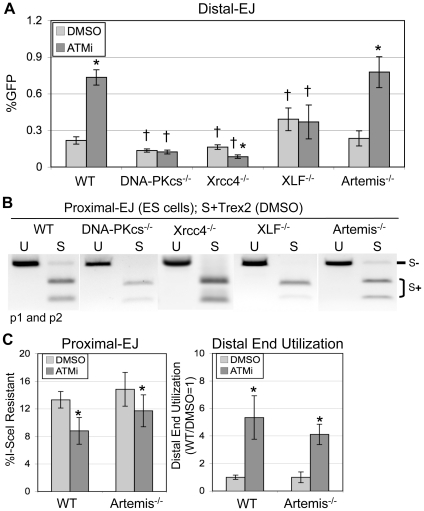
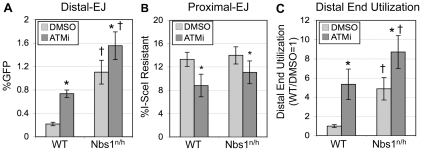
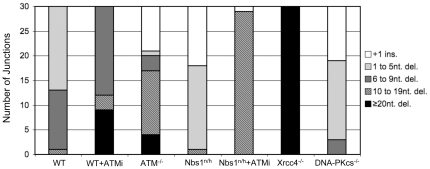
Chromosome rearrangements can form when incorrect ends are matched during end joining (EJ) repair of multiple chromosomal double-strand breaks (DSBs). We tested whether the ATM kinase limits chromosome rearrangements via suppressing incorrect end utilization during EJ repair of multiple DSBs. For this, we developed a system for monitoring EJ of two tandem DSBs that can be repaired using correct ends (Proximal-EJ) or incorrect ends (Distal-EJ, which causes loss of the DNA between the DSBs). In this system, two DSBs are induced in a chromosomal reporter by the meganuclease I-SceI. These DSBs are processed into non-cohesive ends by the exonuclease Trex2, which leads to the formation of I-SceI-resistant EJ products during both Proximal-EJ and Distal-EJ. Using this method, we find that genetic or chemical disruption of ATM causes a substantial increase in Distal-EJ, but not Proximal-EJ. We also find that the increase in Distal-EJ caused by ATM disruption is dependent on classical non-homologous end joining (c-NHEJ) factors, specifically DNA-PKcs, Xrcc4, and XLF. We present evidence that Nbs1-deficiency also causes elevated Distal-EJ, but not Proximal-EJ, to a similar degree as ATM-deficiency. In addition, to evaluate the roles of these factors on end processing, we examined Distal-EJ repair junctions. We found that ATM and Xrcc4 limit the length of deletions, whereas Nbs1 and DNA-PKcs promote short deletions. Thus, the regulation of end processing appears distinct from that of end utilization. In summary, we suggest that ATM is important to limit incorrect end utilization during c-NHEJ.
染色体重排可以在多个染色体双链断裂(DSB)的末端连接(EJ)修复过程中匹配错误的末端时形成。我们测试了 ATM 激酶是否通过在多个 DSB 的 EJ 修复过程中抑制错误末端的利用来限制染色体重排。为此,我们开发了一种监测两个串联 DSB 的 EJ 的系统,这些 DSB 可以使用正确的末端(近端-EJ)或错误的末端(远端-EJ,导致 DSB 之间的 DNA 丢失)进行修复。在这个系统中,通过 meganuclease I-SceI 在染色体报告器中诱导两个 DSB。这些 DSB 通过核酸外切酶 Trex2 处理成非粘性末端,这导致在近端-EJ 和远端-EJ 过程中形成 I-SceI 抗性 EJ 产物。使用这种方法,我们发现 ATM 的遗传或化学破坏会导致远端-EJ 大量增加,但近端-EJ 不会。我们还发现,ATM 破坏引起的远端-EJ 增加依赖于经典非同源末端连接(c-NHEJ)因子,特别是 DNA-PKcs、Xrcc4 和 XLF。我们提供的证据表明,Nbs1 缺陷也会导致类似程度的远端-EJ 增加,但近端-EJ 不会。此外,为了评估这些因素对末端处理的作用,我们检查了远端-EJ 修复连接点。我们发现 ATM 和 Xrcc4 限制了缺失的长度,而 Nbs1 和 DNA-PKcs 促进了短缺失。因此,末端处理的调节似乎与末端利用的调节不同。总之,我们认为 ATM 对于限制 c-NHEJ 过程中的错误末端利用很重要。