Steinmann Institute, University of Bonn, Germany.
Biol Rev Camb Philos Soc. 2011 Feb;86(1):117-55. doi: 10.1111/j.1469-185X.2010.00137.x.
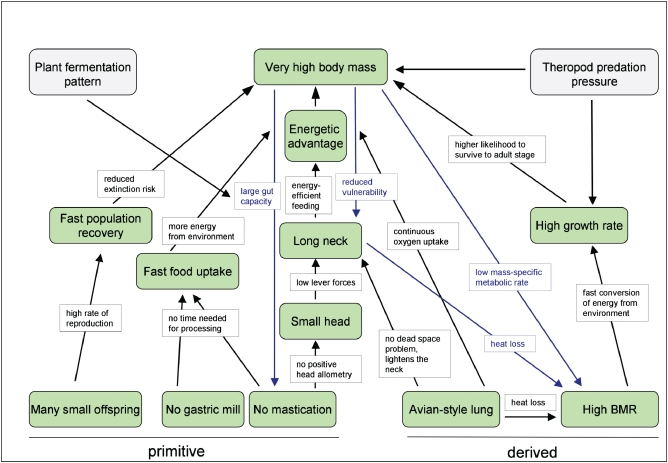
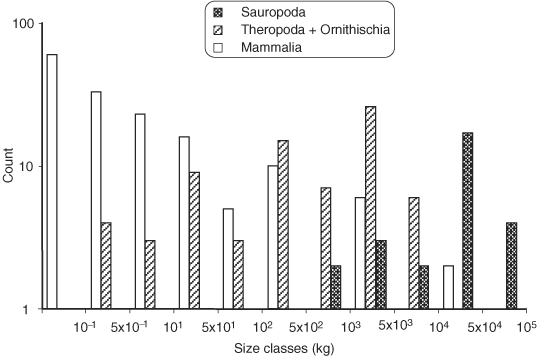
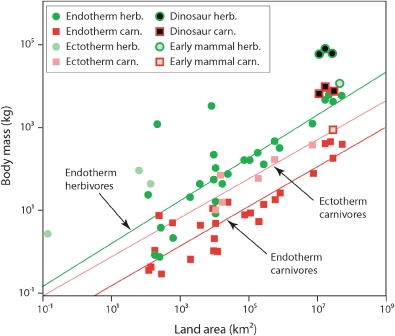
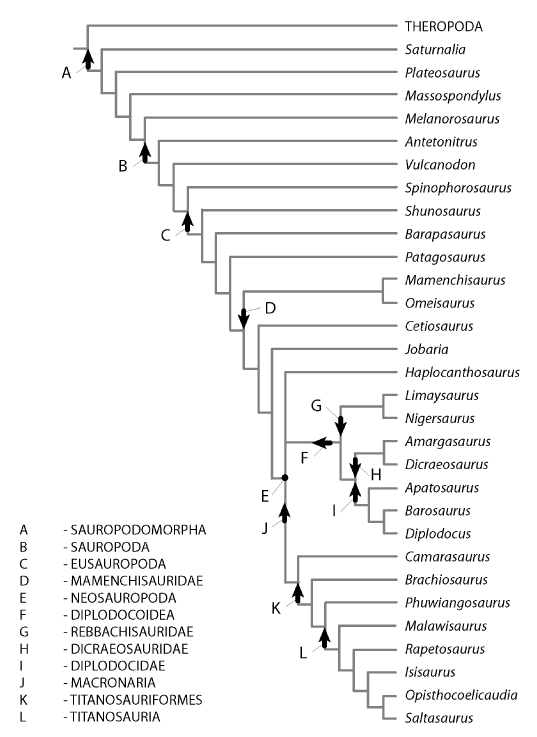
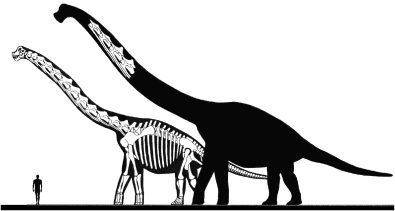
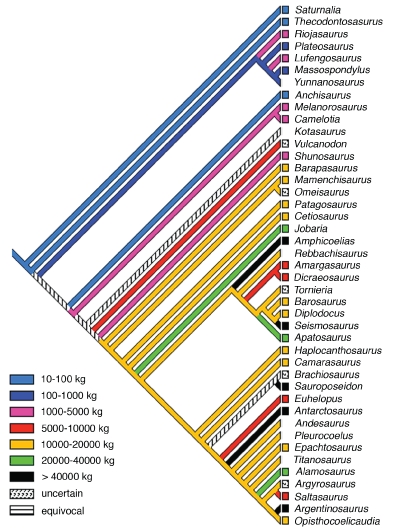
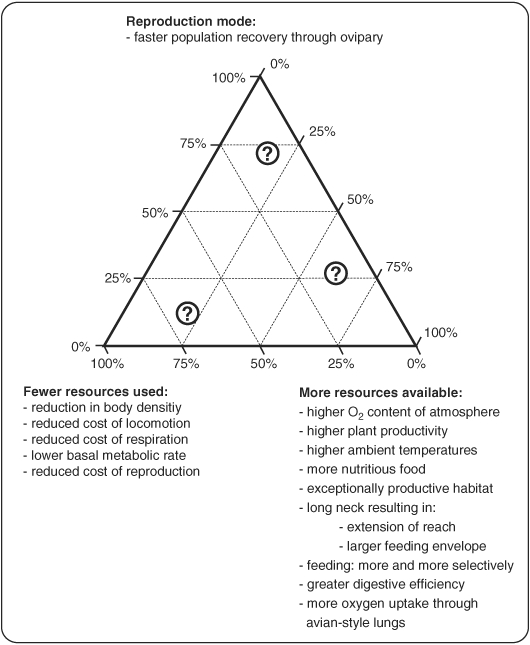
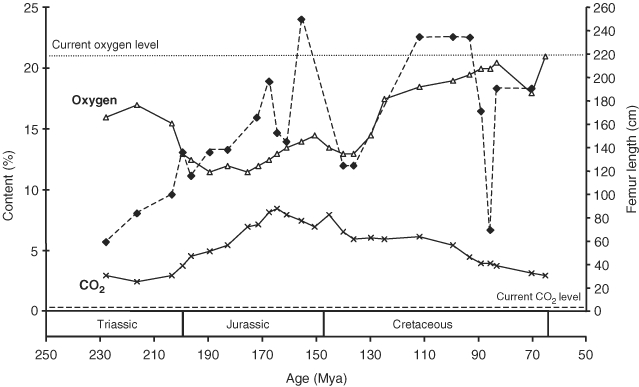
The herbivorous sauropod dinosaurs of the Jurassic and Cretaceous periods were the largest terrestrial animals ever, surpassing the largest herbivorous mammals by an order of magnitude in body mass. Several evolutionary lineages among Sauropoda produced giants with body masses in excess of 50 metric tonnes by conservative estimates. With body mass increase driven by the selective advantages of large body size, animal lineages will increase in body size until they reach the limit determined by the interplay of bauplan, biology, and resource availability. There is no evidence, however, that resource availability and global physicochemical parameters were different enough in the Mesozoic to have led to sauropod gigantism. We review the biology of sauropod dinosaurs in detail and posit that sauropod gigantism was made possible by a specific combination of plesiomorphic characters (phylogenetic heritage) and evolutionary innovations at different levels which triggered a remarkable evolutionary cascade. Of these key innovations, the most important probably was the very long neck, the most conspicuous feature of the sauropod bauplan. Compared to other herbivores, the long neck allowed more efficient food uptake than in other large herbivores by covering a much larger feeding envelope and making food accessible that was out of the reach of other herbivores. Sauropods thus must have been able to take up more energy from their environment than other herbivores. The long neck, in turn, could only evolve because of the small head and the extensive pneumatization of the sauropod axial skeleton, lightening the neck. The small head was possible because food was ingested without mastication. Both mastication and a gastric mill would have limited food uptake rate. Scaling relationships between gastrointestinal tract size and basal metabolic rate (BMR) suggest that sauropods compensated for the lack of particle reduction with long retention times, even at high uptake rates. The extensive pneumatization of the axial skeleton resulted from the evolution of an avian-style respiratory system, presumably at the base of Saurischia. An avian-style respiratory system would also have lowered the cost of breathing, reduced specific gravity, and may have been important in removing excess body heat. Another crucial innovation inherited from basal dinosaurs was a high BMR. This is required for fueling the high growth rate necessary for a multi-tonne animal to survive to reproductive maturity. The retention of the plesiomorphic oviparous mode of reproduction appears to have been critical as well, allowing much faster population recovery than in megaherbivore mammals. Sauropods produced numerous but small offspring each season while land mammals show a negative correlation of reproductive output to body size. This permitted lower population densities in sauropods than in megaherbivore mammals but larger individuals. Our work on sauropod dinosaurs thus informs us about evolutionary limits to body size in other groups of herbivorous terrestrial tetrapods. Ectothermic reptiles are strongly limited by their low BMR, remaining small. Mammals are limited by their extensive mastication and their vivipary, while ornithsichian dinosaurs were only limited by their extensive mastication, having greater average body sizes than mammals.
植食性蜥脚类恐龙生活在侏罗纪和白垩纪时期,是有史以来最大的陆地动物,其体型质量超过了最大的植食性哺乳动物一个数量级。蜥脚形亚目恐龙的几个进化谱系产生了超过 50 公吨的巨型动物,这是保守估计的结果。随着体型增大带来的选择优势,动物谱系的体型将不断增大,直到达到由生物结构、生物学和资源可用性相互作用决定的极限。然而,没有证据表明中生代的资源可用性和全球物理化学参数有足够大的差异,导致了蜥脚类恐龙的巨型化。我们详细回顾了蜥脚类恐龙的生物学,并提出蜥脚类恐龙的巨型化是由特有的祖征(系统发育遗产)和不同层次的进化创新共同作用的结果,引发了显著的进化级联反应。在这些关键创新中,最重要的可能是非常长的脖子,这是蜥脚类恐龙生物结构的最显著特征。与其他食草动物相比,长脖子使它们能够更有效地进食,因为它们的进食范围更大,可以吃到其他食草动物无法触及的食物。因此,蜥脚类恐龙一定能够从环境中获取比其他食草动物更多的能量。反过来,长脖子的进化也只能通过小脑袋和蜥脚类轴向骨骼的广泛充气来实现,这减轻了脖子的重量。小脑袋的形成是因为食物是不经咀嚼就被咽下的。咀嚼和胃磨都会限制食物的摄取率。胃肠道大小与基础代谢率(BMR)之间的比例关系表明,蜥脚类恐龙通过延长食物滞留时间来弥补缺乏颗粒减少的问题,即使在高摄取率的情况下也是如此。轴向骨骼的广泛充气是由鸟类呼吸系统的进化引起的,这种呼吸系统可能起源于蜥脚形亚目恐龙。鸟类呼吸系统还可以降低呼吸成本、减轻比重,并可能在清除多余体热方面发挥重要作用。另一个从基础恐龙继承的关键创新是高基础代谢率(BMR)。这是为了给一个多吨重的动物提供高生长率所必需的燃料。保留原始的卵生繁殖方式似乎也很关键,这使得蜥脚类恐龙的种群恢复速度比巨型食草哺乳动物更快。蜥脚类恐龙每一季都会产下许多但体型较小的后代,而陆地哺乳动物的繁殖输出与体型大小呈负相关。这使得蜥脚类恐龙的种群密度低于巨型食草哺乳动物,但体型更大。我们对蜥脚类恐龙的研究结果为我们了解其他植食性陆地四足动物的体型进化极限提供了信息。冷血爬行动物受限于低基础代谢率(BMR),体型较小。哺乳动物受限于其广泛的咀嚼和胎生,而鸟类恐龙则仅受限于其广泛的咀嚼,其平均体型大于哺乳动物。