Department of Molecular Biosciences, Northwestern University, Evanston, Illinois, USA.
Nat Chem Biol. 2011 Mar;7(3):161-7. doi: 10.1038/nchembio.521. Epub 2011 Jan 30.
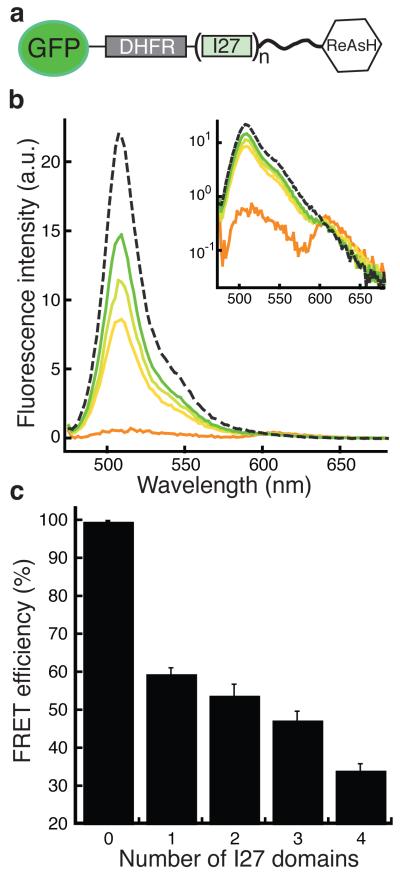
The eukaryotic 26S proteasome controls cellular processes by degrading specific regulatory proteins. Most proteins are targeted for degradation by a signal or degron that consists of two parts: a proteasome-binding tag, typically covalently attached polyubiquitin chains, and an unstructured region that serves as the initiation region for proteasomal proteolysis. Here we have characterized how the arrangement of the two degron parts in a protein affects degradation. We found that a substrate is degraded efficiently only when its initiation region is of a certain minimal length and is appropriately separated in space from the proteasome-binding tag. Regions that are located too close or too far from the proteasome-binding tag cannot access the proteasome and induce degradation. These spacing requirements are different for a polyubiquitin chain and a ubiquitin-like domain. Thus, the arrangement and location of the proteasome initiation region affect a protein's fate and are important in selecting proteins for proteasome-mediated degradation.
真核生物 26S 蛋白酶体通过降解特定的调节蛋白来控制细胞过程。大多数蛋白质的靶向降解是由一个信号或降解元件(degron)组成的,该信号由两部分组成:一个蛋白酶体结合标签,通常是共价连接的多聚泛素链,和一个作为蛋白酶体蛋白水解起始区域的无结构区域。在这里,我们描述了蛋白质中两个降解元件部分的排列如何影响降解。我们发现,只有当起始区域具有一定的最小长度并且与蛋白酶体结合标签在空间上适当分离时,底物才能有效地被降解。与蛋白酶体结合标签太近或太远的区域无法进入蛋白酶体并诱导降解。对于多聚泛素链和泛素样结构域,这些间隔要求是不同的。因此,蛋白酶体起始区域的排列和位置影响蛋白质的命运,并且在选择蛋白质进行蛋白酶体介导的降解中很重要。