Biotechnology, Dept of Chemistry, Lund University, P,O, Box 124, SE-221 00 Lund, Sweden.
BMC Biochem. 2011 Feb 23;12:11. doi: 10.1186/1471-2091-12-11.
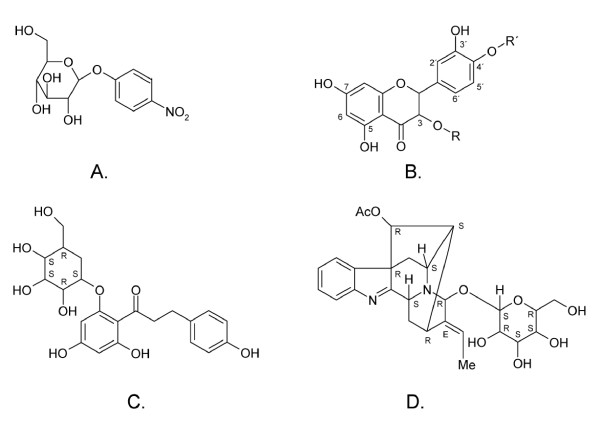
The thermostable β-glucosidase (TnBgl1A) from Thermotoga neapolitana is a promising biocatalyst for hydrolysis of glucosylated flavonoids and can be coupled to extraction methods using pressurized hot water. Hydrolysis has however been shown to be dependent on the position of the glucosylation on the flavonoid, and e.g. quercetin-3-glucoside (Q3) was hydrolysed slowly. A set of mutants of TnBgl1A were thus created to analyse the influence on the kinetic parameters using the model substrate para-nitrophenyl-β-D-glucopyranoside (pNPGlc), and screened for hydrolysis of Q3.
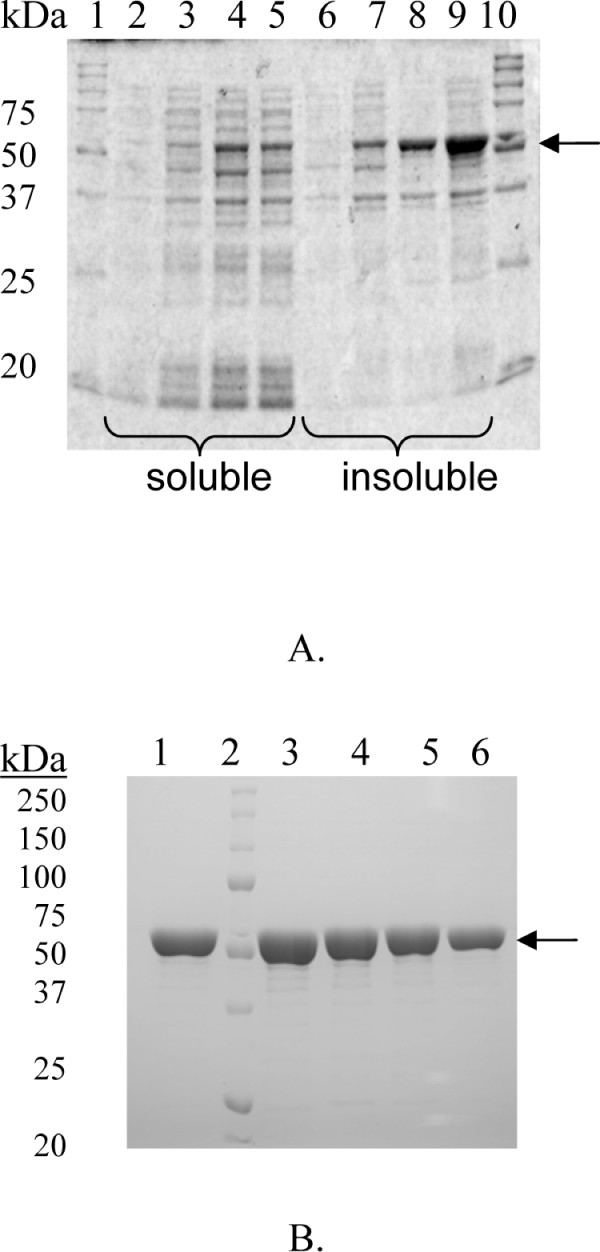
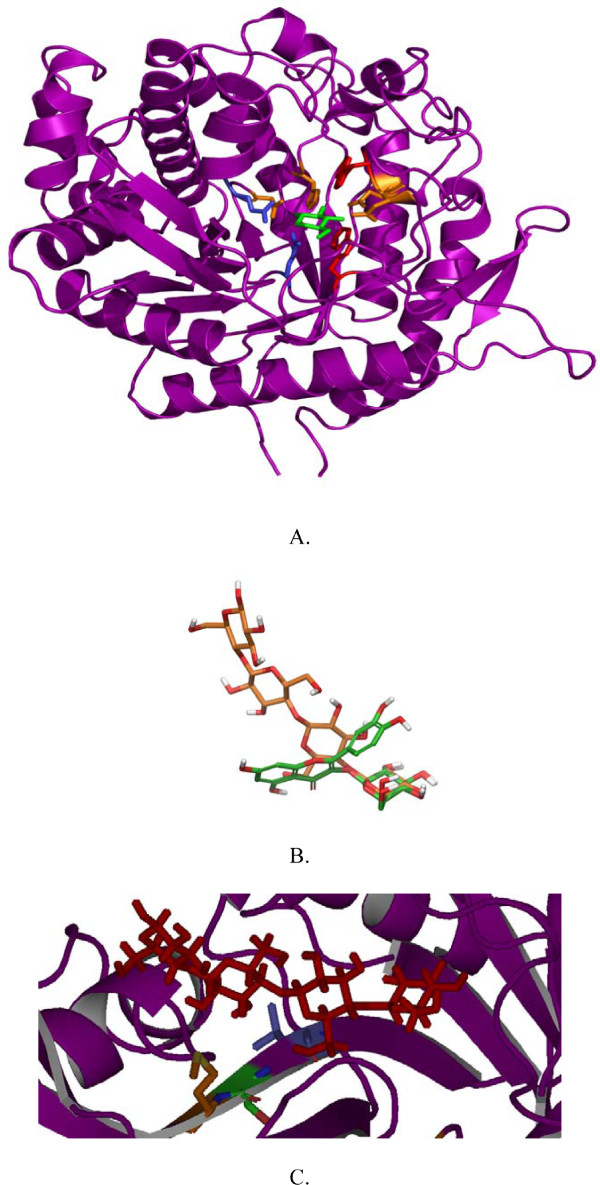
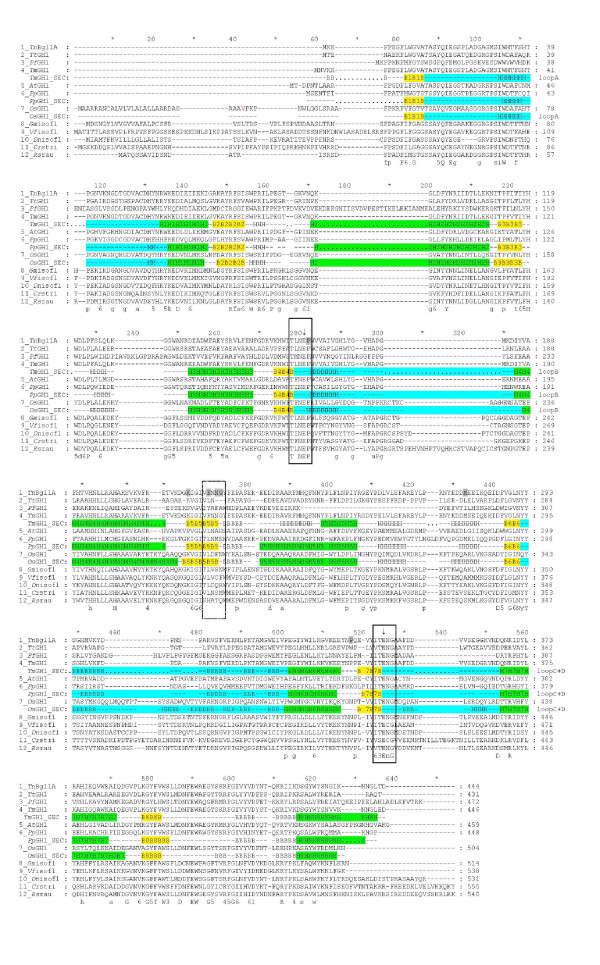
Structural analysis pinpointed an area in the active site pocket with non-conserved residues between specificity groups in glycoside hydrolase family 1 (GH1). Three residues in this area located on β-strand 5 (F219, N221, and G222) close to sugar binding sub-site +2 were selected for mutagenesis and amplified in a protocol that introduced a few spontaneous mutations. Eight mutants (four triple: F219L/P165L/M278I, N221S/P165L/M278I, G222Q/P165L/M278I, G222Q/V203M/K214R, two double: F219L/K214R, N221S/P342L and two single: G222M and N221S) were produced in E. coli, and purified to apparent homogeneity. Thermostability, measured as Tm by differential scanning calorimetry (101.9°C for wt), was kept in the mutated variants and significant decrease (ΔT of 5-10°C) was only observed for the triple mutants. The exchanged residue(s) in the respective mutant resulted in variations in KM and turnover. The KM-value was only changed in variants mutated at position 221 (N221S) and was in all cases monitored as a 2-3 × increase for pNPGlc, while the KM decreased a corresponding extent for Q3.Turnover was only significantly changed using pNPGlc, and was decreased 2-3 × in variants mutated at position 222, while the single, double and triple mutated variants carrying a mutation at position 221 (N221S) increased turnover up to 3.5 × compared to the wild type. Modelling showed that the mutation at position 221, may alter the position of N291 resulting in increased hydrogen bonding of Q3 (at a position corresponding to the +1 subsite) which may explain the decrease in KM for this substrate.
These results show that residues at the +2 subsite are interesting targets for mutagenesis and mutations at these positions can directly or indirectly affect both KM and turnover. An affinity change, leading to a decreased KM, can be explained by an altered position of N291, while the changes in turnover are more difficult to explain and may be the result of smaller conformational changes in the active site.
来自嗜热栖热菌(Thermotoga neapolitana)的耐热β-葡萄糖苷酶(TnBgl1A)是水解葡萄糖基化黄酮类化合物的有前途的生物催化剂,并且可以与使用加压热水的提取方法结合使用。然而,水解已被证明取决于黄酮类化合物上的葡糖基化位置,例如,槲皮素-3-葡萄糖苷(Q3)的水解速度较慢。因此,创建了一组 TnBgl1A 的突变体,以使用模型底物对硝基苯-β-D-吡喃葡萄糖苷(pNPGlc)分析对动力学参数的影响,并筛选 Q3 的水解。
结构分析指出糖苷水解酶家族 1(GH1)中特异性基团之间的活性位点口袋中的一个区域存在非保守残基。该区域中位于β-链 5 上的三个残基(F219、N221 和 G222)靠近糖结合亚位点+2,被选择进行诱变,并在引入一些自发突变的方案中进行扩增。产生了 8 种突变体(4 种三重突变体:F219L/P165L/M278I、N221S/P165L/M278I、G222Q/P165L/M278I、G222Q/V203M/K214R,2 种双突变体:F219L/K214R、N221S/P342L 和 2 种单突变体:G222M 和 N221S),在大肠杆菌中产生,并纯化至明显均一。通过差示扫描量热法(wt 的 Tm 为 101.9°C)测量的热稳定性在突变变体中得以保留,仅在三重突变体中观察到显著降低(5-10°C 的ΔT)。相应突变体中交换的残基导致 KM 和周转率的变化。仅在突变位置为 221 的变体(N221S)中改变了 KM 值,并且在所有情况下,pNPGlc 的 KM 值均监测为 2-3×增加,而对于 Q3 的 KM 值相应降低。仅使用 pNPGlc 显著改变了周转率,而在突变位置为 222 的变体中降低了 2-3×,而在突变位置为 221(N221S)的单突变体、双突变体和三突变体的周转率与野生型相比增加了 3.5 倍。建模表明,位置 221 的突变可能改变 N291 的位置,导致 Q3(在对应于+1亚位点的位置)的氢键增加,这可以解释该底物 KM 的降低。
这些结果表明+2 亚位点的残基是诱变的有趣目标,并且这些位置的突变可以直接或间接影响 KM 和周转率。亲和力的变化导致 KM 降低,可以通过 N291 的位置改变来解释,而周转率的变化更难解释,可能是由于活性位点的较小构象变化所致。