Department of Biology, Carolina Center for Genome Sciences and Lineberger Comprehensive Cancer Center, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina, United States of America.
PLoS One. 2011 Apr 28;6(4):e19060. doi: 10.1371/journal.pone.0019060.
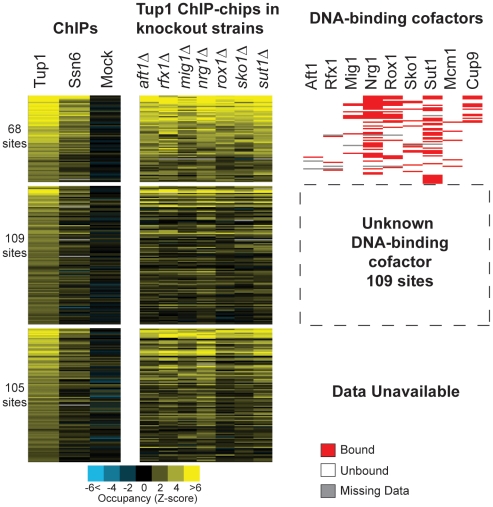
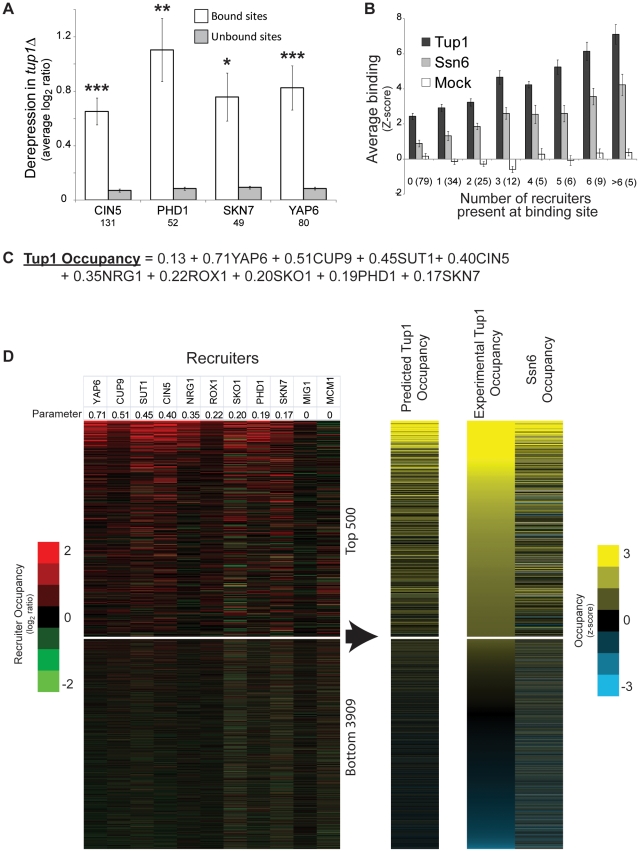
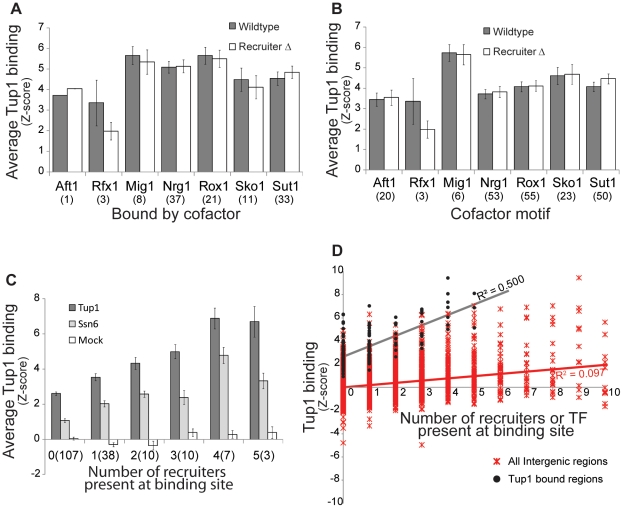
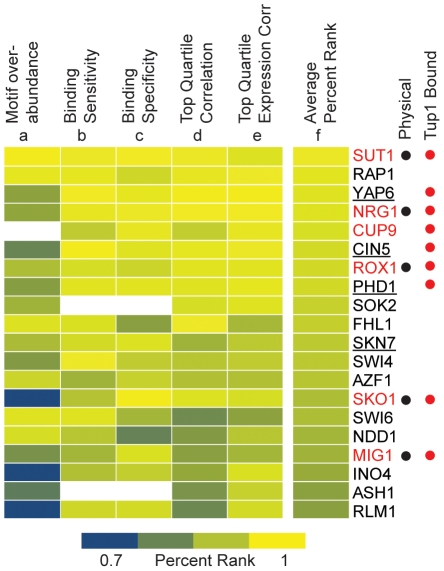
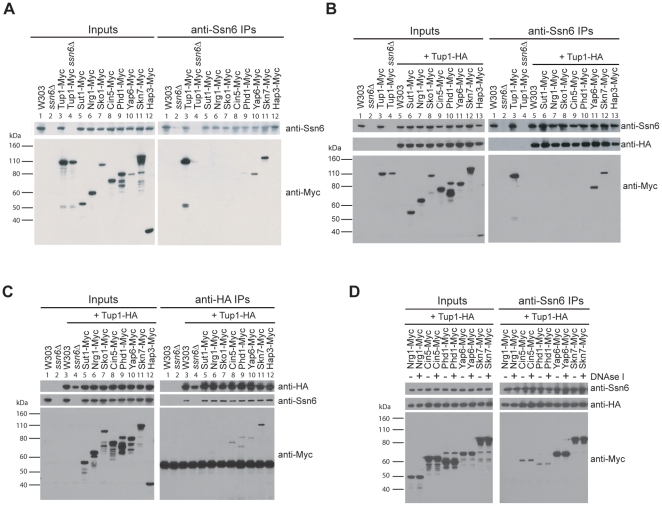
Maintaining the proper expression of the transcriptome during development or in response to a changing environment requires a delicate balance between transcriptional regulators with activating and repressing functions. The budding yeast transcriptional co-repressor Tup1-Ssn6 is a model for studying similar repressor complexes in multicellular eukaryotes. Tup1-Ssn6 does not bind DNA directly, but is directed to individual promoters by one or more DNA-binding proteins, referred to as Tup1 recruiters. This functional architecture allows the Tup1-Ssn6 to modulate the expression of genes required for the response to a variety of cellular stresses. To understand the targeting or the Tup1-Ssn6 complex, we determined the genomic distribution of Tup1 and Ssn6 by ChIP-chip. We found that most loci bound by Tup1-Ssn6 could not be explained by co-occupancy with a known recruiting cofactor and that deletion of individual known Tup1 recruiters did not significantly alter the Tup1 binding profile. These observations suggest that new Tup1 recruiting proteins remain to be discovered and that Tup1 recruitment typically depends on multiple recruiting cofactors. To identify new recruiting proteins, we computationally screened for factors with binding patterns similar to the observed Tup1-Ssn6 genomic distribution. Four top candidates, Cin5, Skn7, Phd1, and Yap6, all known to be associated with stress response gene regulation, were experimentally confirmed to physically interact with Tup1 and/or Ssn6. Incorporating these new recruitment cofactors with previously characterized cofactors now explains the majority of Tup1 targeting across the genome, and expands our understanding of the mechanism by which Tup1-Ssn6 is directed to its targets.
在发育过程中或应对环境变化时,维持转录组的适当表达需要具有激活和抑制功能的转录调节剂之间的微妙平衡。芽殖酵母转录共抑制因子 Tup1-Ssn6 是研究多细胞真核生物中类似的抑制复合物的模型。Tup1-Ssn6 不直接结合 DNA,而是由一个或多个 DNA 结合蛋白(称为 Tup1 招募因子)定向到单个启动子。这种功能结构允许 Tup1-Ssn6 调节对各种细胞应激反应所需基因的表达。为了了解 Tup1-Ssn6 复合物的靶向作用,我们通过 ChIP-chip 确定了 Tup1 和 Ssn6 的基因组分布。我们发现,Tup1-Ssn6 结合的大多数位点不能用与已知招募共因子的共占据来解释,并且删除单个已知的 Tup1 招募因子不会显著改变 Tup1 结合谱。这些观察结果表明,仍然需要发现新的 Tup1 招募蛋白,并且 Tup1 的招募通常依赖于多个招募共因子。为了鉴定新的招募蛋白,我们通过计算筛选出与观察到的 Tup1-Ssn6 基因组分布具有相似结合模式的因子。Cin5、Skn7、Phd1 和 Yap6 这四个候选因子,均已知与应激反应基因调控有关,它们均被实验证实与 Tup1 和/或 Ssn6 物理相互作用。将这些新的招募共因子与以前表征的共因子结合起来,现在可以解释基因组中大多数 Tup1 靶向作用的原因,并扩展了我们对 Tup1-Ssn6 定向到其靶标的机制的理解。