Developmental Biology Program, Memorial Sloan-Kettering Cancer Center, New York, New York, USA.
PLoS Genet. 2011 Jun;7(6):e1002080. doi: 10.1371/journal.pgen.1002080. Epub 2011 Jun 2.
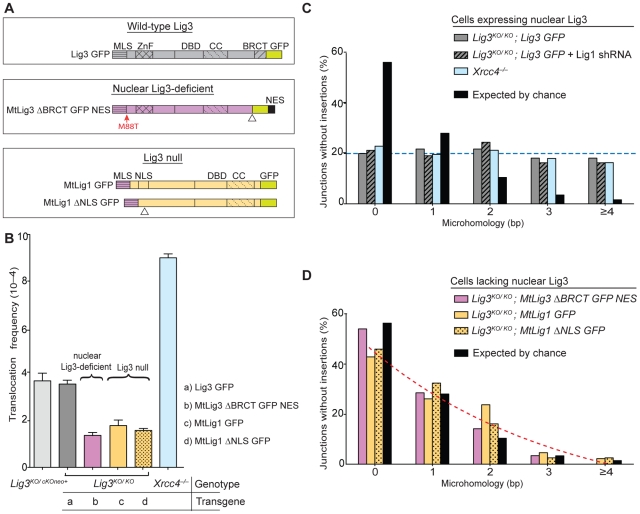
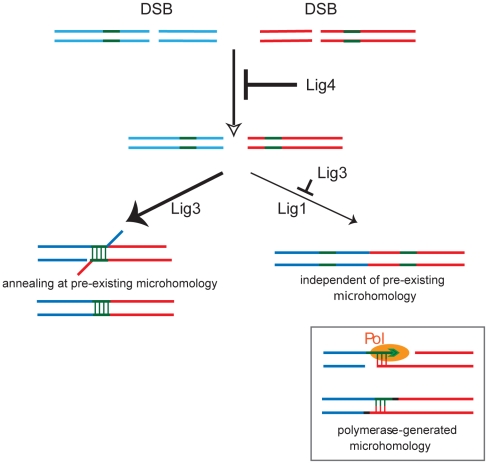
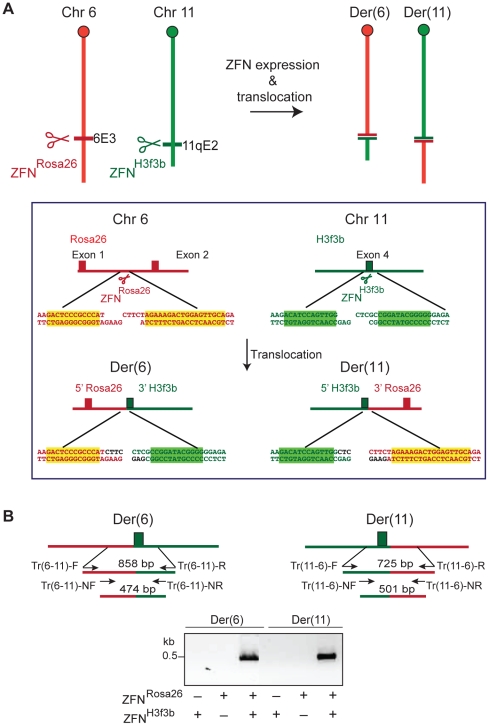
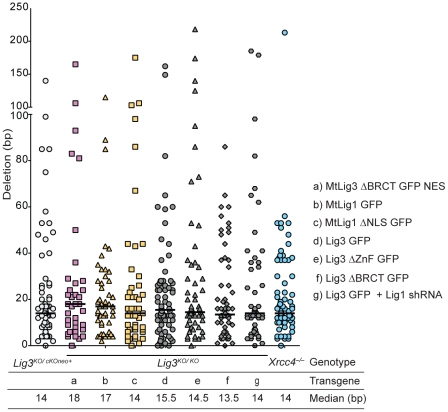
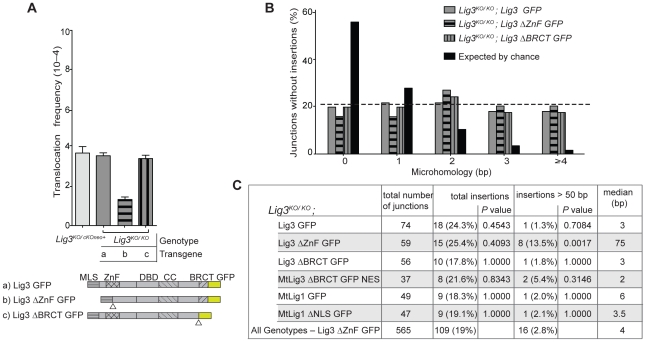
Nonhomologous end-joining (NHEJ) is the primary DNA repair pathway thought to underlie chromosomal translocations and other genomic rearrangements in somatic cells. The canonical NHEJ pathway, including DNA ligase IV (Lig4), suppresses genomic instability and chromosomal translocations, leading to the notion that a poorly defined, alternative NHEJ (alt-NHEJ) pathway generates these rearrangements. Here, we investigate the DNA ligase requirement of chromosomal translocation formation in mouse cells. Mammals have two other DNA ligases, Lig1 and Lig3, in addition to Lig4. As deletion of Lig3 results in cellular lethality due to its requirement in mitochondria, we used recently developed cell lines deficient in nuclear Lig3 but rescued for mitochondrial DNA ligase activity. Further, zinc finger endonucleases were used to generate DNA breaks at endogenous loci to induce translocations. Unlike with Lig4 deficiency, which causes an increase in translocation frequency, translocations are reduced in frequency in the absence of Lig3. Residual translocations in Lig3-deficient cells do not show a bias toward use of pre-existing microhomology at the breakpoint junctions, unlike either wild-type or Lig4-deficient cells, consistent with the notion that alt-NHEJ is impaired with Lig3 loss. By contrast, Lig1 depletion in otherwise wild-type cells does not reduce translocations or affect microhomology use. However, translocations are further reduced in Lig3-deficient cells upon Lig1 knockdown, suggesting the existence of two alt-NHEJ pathways, one that is biased toward microhomology use and requires Lig3 and a back-up pathway which does not depend on microhomology and utilizes Lig1.
非同源末端连接(NHEJ)是一种主要的 DNA 修复途径,被认为是体细胞中染色体易位和其他基因组重排的基础。经典的 NHEJ 途径,包括 DNA 连接酶 IV(Lig4),抑制基因组不稳定性和染色体易位,导致出现了一种定义不明确的替代 NHEJ(alt-NHEJ)途径产生这些重排的观点。在这里,我们研究了 DNA 连接酶在小鼠细胞中染色体易位形成中的需求。哺乳动物除了 Lig4 之外,还有另外两种 DNA 连接酶 Lig1 和 Lig3。由于 Lig3 在线粒体中的缺失会导致细胞死亡,因此我们使用了最近开发的缺乏核 Lig3 但能挽救线粒体 DNA 连接酶活性的细胞系。此外,锌指内切酶被用来在内源性基因座上产生 DNA 断裂,以诱导易位。与 Lig4 缺失导致易位频率增加不同,Lig3 缺失导致易位频率降低。与野生型或 Lig4 缺失细胞不同,Lig3 缺失细胞中的残留易位在断裂点连接处不倾向于使用预先存在的微同源性,这与 alt-NHEJ 在 Lig3 缺失时受损的观点一致。相比之下,在其他方面为野生型的细胞中 Lig1 的耗竭不会减少易位或影响微同源性的使用。然而,在 Lig3 缺失细胞中 Lig1 敲低后,易位进一步减少,这表明存在两种 alt-NHEJ 途径,一种偏向于微同源性的使用,需要 Lig3,另一种备用途径不依赖于微同源性,利用 Lig1。