Department of Genetics, Evolution and Environment, University College London, London, UK.
Biol Direct. 2011 Jun 30;6:35. doi: 10.1186/1745-6150-6-35.
All complex life on Earth is eukaryotic. All eukaryotic cells share a common ancestor that arose just once in four billion years of evolution. Prokaryotes show no tendency to evolve greater morphological complexity, despite their metabolic virtuosity. Here I argue that the eukaryotic cell originated in a unique prokaryotic endosymbiosis, a singular event that transformed the selection pressures acting on both host and endosymbiont.
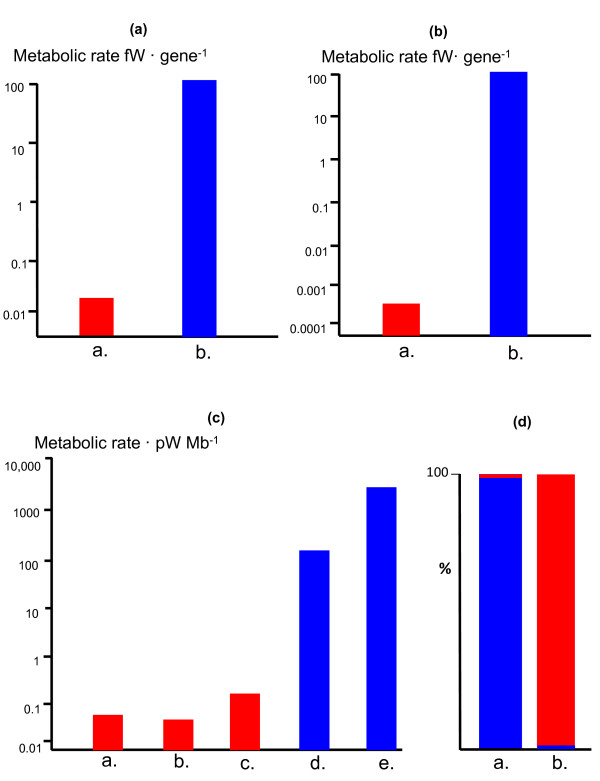
The reductive evolution and specialisation of endosymbionts to mitochondria resulted in an extreme genomic asymmetry, in which the residual mitochondrial genomes enabled the expansion of bioenergetic membranes over several orders of magnitude, overcoming the energetic constraints on prokaryotic genome size, and permitting the host cell genome to expand (in principle) over 200,000-fold. This energetic transformation was permissive, not prescriptive; I suggest that the actual increase in early eukaryotic genome size was driven by a heavy early bombardment of genes and introns from the endosymbiont to the host cell, producing a high mutation rate. Unlike prokaryotes, with lower mutation rates and heavy selection pressure to lose genes, early eukaryotes without genome-size limitations could mask mutations by cell fusion and genome duplication, as in allopolyploidy, giving rise to a proto-sexual cell cycle. The side effect was that a large number of shared eukaryotic basal traits accumulated in the same population, a sexual eukaryotic common ancestor, radically different to any known prokaryote.
The combination of massive bioenergetic expansion, release from genome-size constraints, and high mutation rate favoured a protosexual cell cycle and the accumulation of eukaryotic traits. These factors explain the unique origin of eukaryotes, the absence of true evolutionary intermediates, and the evolution of sex in eukaryotes but not prokaryotes.
This article was reviewed by: Eugene Koonin, William Martin, Ford Doolittle and Mark van der Giezen. For complete reports see the Reviewers' Comments section.
地球上所有复杂的生命都是真核生物。所有真核细胞都拥有一个共同的祖先,这个祖先是在 40 亿年的进化过程中仅出现过一次的。尽管原核生物具有代谢灵活性,但它们没有进化出更高形态复杂性的趋势。在这里,我认为真核细胞起源于独特的原核内共生体,这一独特的事件改变了宿主和内共生体所面临的选择压力。
内共生体向线粒体的还原进化和特化导致了基因组的极度不对称,其中残余的线粒体基因组使生物能膜扩展了几个数量级,克服了原核生物基因组大小的能量限制,并允许宿主细胞基因组(原则上)扩展 200000 倍以上。这种能量转换是允许的,而不是规定的;我认为,早期真核生物基因组大小的实际增加是由内共生体向宿主细胞大量基因和内含子的冲击所驱动的,从而产生了高突变率。与突变率较低且有强烈基因丢失选择压力的原核生物不同,早期真核生物没有基因组大小的限制,可以通过细胞融合和基因组复制来掩盖突变,就像异源多倍体一样,从而产生原始的有性细胞周期。其副作用是,大量共同的真核生物基础特征在同一群体中积累,形成一个有性真核生物的共同祖先,与任何已知的原核生物都有很大的不同。
大规模生物能扩张、摆脱基因组大小限制以及高突变率的结合,有利于原始有性细胞周期和真核生物特征的积累。这些因素解释了真核生物的独特起源、真正进化中间产物的缺失,以及真核生物而非原核生物中性的进化。
本文由 Eugene Koonin、William Martin、Ford Doolittle 和 Mark van der Giezen 进行了评审。完整的评审报告请参见“评论者意见”部分。