Maurice Wilkins Centre for Molecular Biodiscovery and School of Biological Sciences, University of Auckland, Auckland, New Zealand.
PLoS One. 2011;6(7):e22095. doi: 10.1371/journal.pone.0022095. Epub 2011 Jul 8.
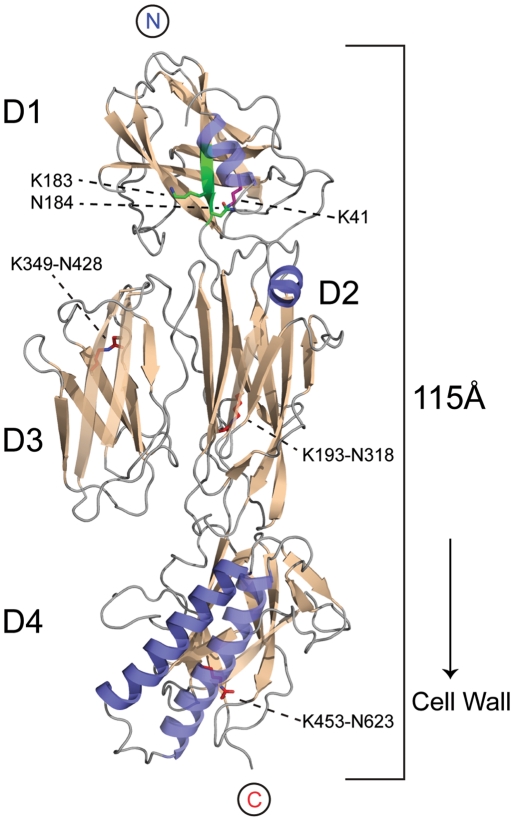
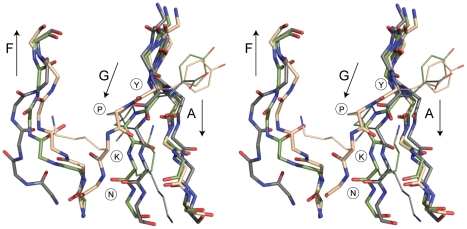
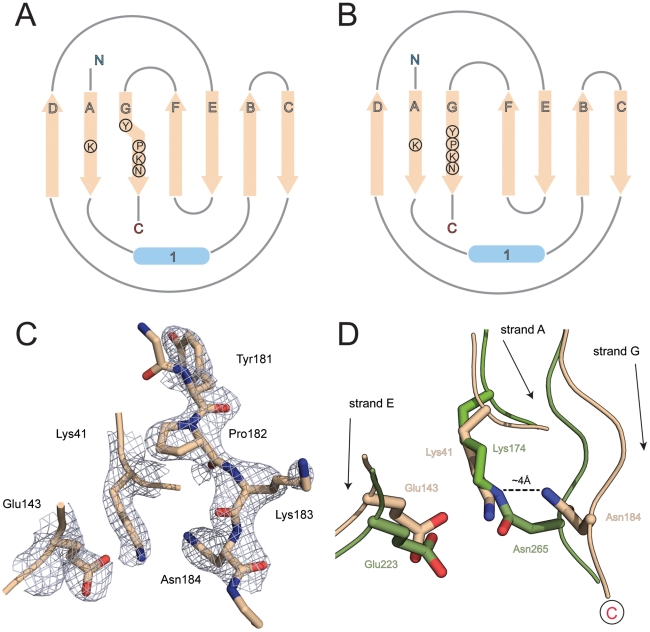
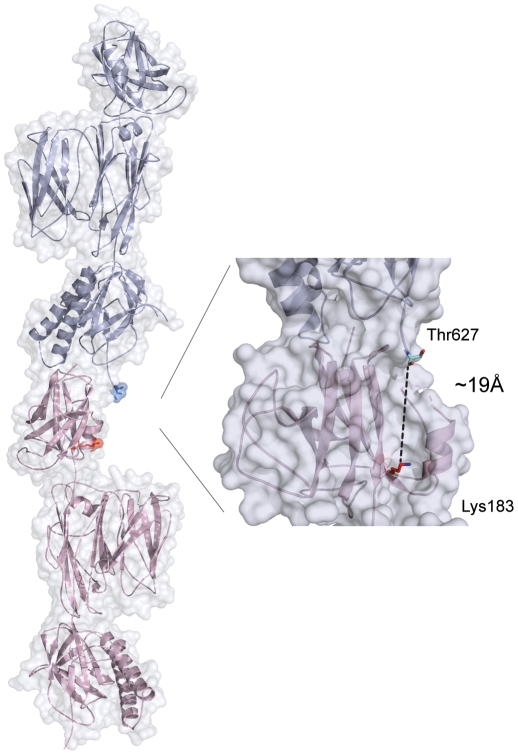
The surface of the pneumococcal cell is adorned with virulence factors including pili. The major pilin RrgB, which forms the pilus shaft on pathogenic Streptococcus pneumoniae, comprises four immunoglobulin (Ig)-like domains, each with a common CnaB topology. The three C-terminal domains are each stabilized by internal Lys-Asn isopeptide bonds, formed autocatalytically with the aid of an essential Glu residue. The structure and orientation of the crucial N-terminal domain, which provides the covalent linkage to the next pilin subunit in the shaft, however, remain incompletely characterised. We report the crystal structure of full length RrgB, solved by X-ray crystallography at 2.8 Å resolution. The N-terminal (D1) domain makes few contacts with the rest of the RrgB structure, and has higher B-factors. This may explain why D1 is readily lost by proteolysis, as are the N-terminal domains of many major pilins. D1 is also found to have a triad of Lys, Asn and Glu residues in the same topological positions as in the other domains, yet mass spectrometry and the crystal structure show that no internal isopeptide bond is formed. We show that this is because β-strand G of D1, which carries the Asn residue, diverges from β-strand A, carrying the Lys residue, such that these residues are too far apart for bond formation. Strand G also carries the YPKN motif that provides the essential Lys residue for the sortase-mediated intermolecular linkages along the pilus shaft. Interaction with the sortase and formation of the intermolecular linkage could result in a change in the orientation of this strand, explaining why isopeptide bond formation in the N-terminal domains of some major pilins appears to take place only upon assembly of the pili.
肺炎球菌细胞表面装饰有包括菌毛在内的毒力因子。主要的 Pilin RrgB 形成致病性肺炎链球菌的菌毛轴,由四个免疫球蛋白(Ig)样结构域组成,每个结构域都具有共同的 CnaB 拓扑结构。三个 C 端结构域每个都被内部 Lys-Asn 异肽键稳定,该键在必需的 Glu 残基的辅助下自动形成。然而,对于提供轴中与下一个菌毛亚基共价连接的关键 N 端结构域的结构和取向,仍不完全清楚。我们通过 X 射线晶体学以 2.8Å 的分辨率解析了全长 RrgB 的晶体结构。N 端(D1)结构域与 RrgB 结构的其余部分很少接触,并且具有较高的 B 因子。这可能解释了为什么 D1 很容易被蛋白酶水解,许多主要菌毛的 N 端结构域也是如此。还发现 D1 具有 Lys、Asn 和 Glu 残基三联体,其拓扑位置与其他结构域相同,但质谱和晶体结构表明没有形成内部异肽键。我们表明,这是因为 D1 的β-链 G 携带 Asn 残基,与携带 Lys 残基的β-链 A 发散,使得这些残基相距太远无法形成键。链 G 还携带 YPKN 基序,该基序为沿菌毛轴的天冬酰胺酶介导的分子间连接提供必需的 Lys 残基。与天冬酰胺酶的相互作用和分子间连接的形成可能导致该链的取向发生变化,这解释了为什么某些主要菌毛的 N 端结构域中的异肽键形成似乎仅在菌毛组装时才发生。