Organismic Botany, Institute of Evolution and Ecology, University of Tübingen, Tübingen, Germany.
BMC Evol Biol. 2011 Jul 19;11:213. doi: 10.1186/1471-2148-11-213.
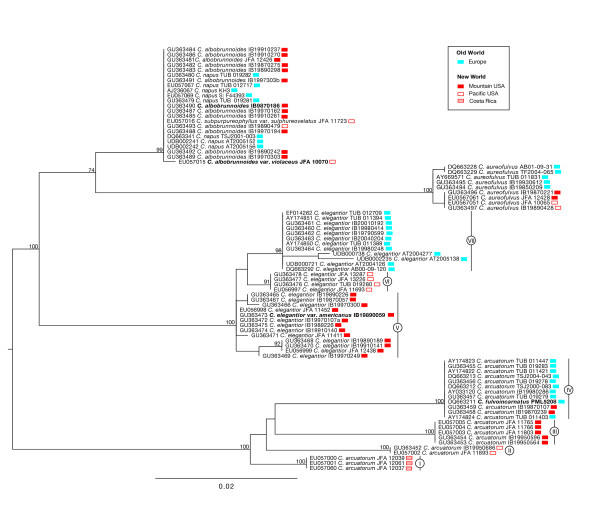
Cortinarius species in section Calochroi display local, clinal and circumboreal patterns of distribution across the Northern Hemisphere where these ectomycorrhizal fungi occur with host trees throughout their geographical range within a continent, or have disjunct intercontinental distributions, the origins of which are not understood. We inferred evolutionary histories of four species, 1) C. arcuatorum, 2) C. aureofulvus, 3) C. elegantior and 4) C. napus, from populations distributed throughout the Old World, and portions of the New World (Central- and North America) based on genetic variation of 154 haplotype internal transcribed spacer (ITS) sequences from 83 population samples. By describing the population structure of these species across their geographical distribution, we attempt to identify their historical migration and patterns of diversification.
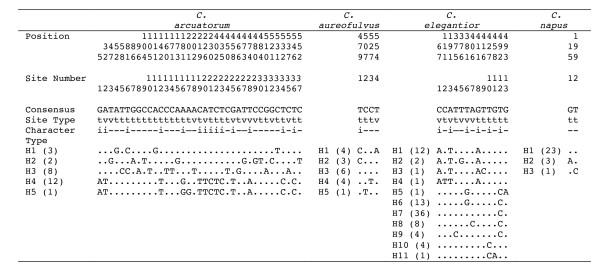
Models of population structure from nested clade, demographic and coalescent-based analyses revealed genetically differentiated and geographically structured haplotypes in C. arcuatorum and C. elegantior, while C. aureofulvus showed considerably less population structure and C. napus lacked sufficient genetic differentiation to resolve any population structure. Disjunct populations within C. arcuatorum, C. aureofulvus and C. elegantior show little or no morphological differentiation, whereas in C. napus there is a high level of homoplasy and phenotypic plasticity for veil and lamellae colour. The ITS sequences of the type specimens of C. albobrunnoides and C. albobrunnoides var. violaceovelatus were identical to one another and are treated as one species with a wider range of geographic distribution under C. napus.
Our results indicate that each of the Calochroi species has undergone a relatively independent evolutionary history, hypothesised as follows: 1) a widely distributed ancestral population of C. arcuatorum diverged into distinctive sympatric populations in the New World; 2) two divergent lineages in C. elegantior gave rise to the New World and Old World haplotypes, respectively; and 3) the low levels of genetic divergence within C. aureofulvus and C. napus may be the result of more recent demographic population expansions. The scenario of migration via the Bering Land Bridge provides the most probable explanation for contemporaneous disjunct geographic distributions of these species, but it does not offer an explanation for the low degree of genetic divergence between populations of C. aureofulvus and C. napus. Our findings are mostly consistent with the designation of New World allopatric populations as separate species from the European counterpart species C. arcuatorum and C. elegantior. We propose the synonymy of C. albobrunnoides, C. albobrunnoides var. violaceovelatus and C. subpurpureophyllus var. sulphureovelatus with C. napus. The results also reinforce previous observations that linked C. arcuatorum and C. aureofulvus displaying distributions in parts of North America and Europe. Interpretations of the population structure of these fungi suggest that host tree history has heavily influenced their modern distributions; however, the complex issues related to co-migration of these fungi with their tree hosts remain unclear at this time.

Calochroi 节中的 Cortinarius 物种在北半球具有局部、渐变和环绕北极的分布模式,这些外生菌根真菌在其地理范围内与宿主树一起出现,或者具有离散的洲际分布,其起源尚不清楚。我们根据来自 83 个种群样本的 154 个单倍型内部转录间隔区(ITS)序列的遗传变异,推断了分布于旧世界和新大陆(中美洲和北美洲)部分地区的四个物种的进化历史,1)C. arcuatorum,2)C. aureofulvus,3)C. elegantior 和 4)C. napus。通过描述这些物种在其地理分布范围内的种群结构,我们试图确定它们的历史迁移和多样化模式。
嵌套进化枝、人口统计学和基于合并的分析模型揭示了 C. arcuatorum 和 C. elegantior 中具有遗传分化和地理结构的单倍型,而 C. aureofulvus 显示出相当少的种群结构,并且 C. napus 缺乏足够的遗传分化来解决任何种群结构问题。C. arcuatorum、C. aureofulvus 和 C. elegantior 中的离散种群表现出很少或没有形态分化,而在 C. napus 中,帽和菌褶颜色存在高度的同形性和表型可塑性。C. albobrunnoides 和 C. albobrunnoides var. violaceovelatus 的模式标本的 ITS 序列彼此相同,并且在 C. napus 下被视为具有更广泛地理分布范围的一个物种。
我们的结果表明,每个 Calochroi 物种都经历了相对独立的进化历史,假设如下:1)一个广泛分布的祖先种群 C. arcuatorum 分化为新大陆的特有种群;2)C. elegantior 的两个分歧谱系分别产生了新大陆和旧世界的单倍型;3)C. aureofulvus 和 C. napus 内的遗传分化水平较低可能是最近人口扩张的结果。通过白令陆桥迁移的情景为这些物种同时发生的离散地理分布提供了最可能的解释,但它并没有解释 C. aureofulvus 和 C. napus 种群之间遗传分化程度低的原因。我们的发现与新大陆的隔离种群被指定为与欧洲对应种 C. arcuatorum 和 C. elegantior 不同的物种的观点基本一致。我们建议将 C. albobrunnoides、C. albobrunnoides var. violaceovelatus 和 C. subpurpureophyllus var. sulphureovelatus 与 C. napus 同义。这些结果也进一步证实了之前的观察结果,即 C. arcuatorum 和 C. aureofulvus 显示分布在北美洲和欧洲部分地区。对这些真菌种群结构的解释表明,宿主树历史对其现代分布有很大影响;然而,目前尚不清楚这些真菌与它们的宿主树共同迁移的复杂问题。