Division of Medicinal Chemistry, Leiden/Amsterdam Centre for Drug Research, Leiden University, P.O. Box 9502, 2300, Leiden, The Netherlands.
Purinergic Signal. 2012 Mar;8(1):23-38. doi: 10.1007/s11302-011-9251-x. Epub 2011 Aug 5.
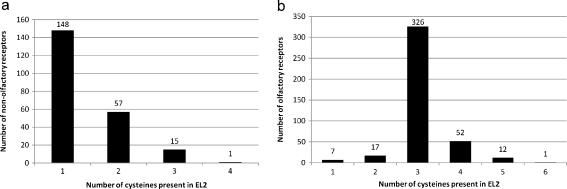
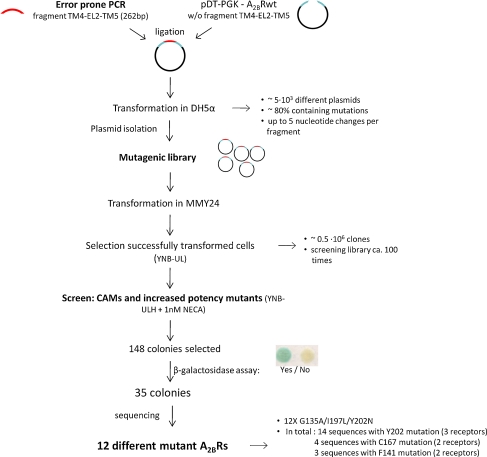
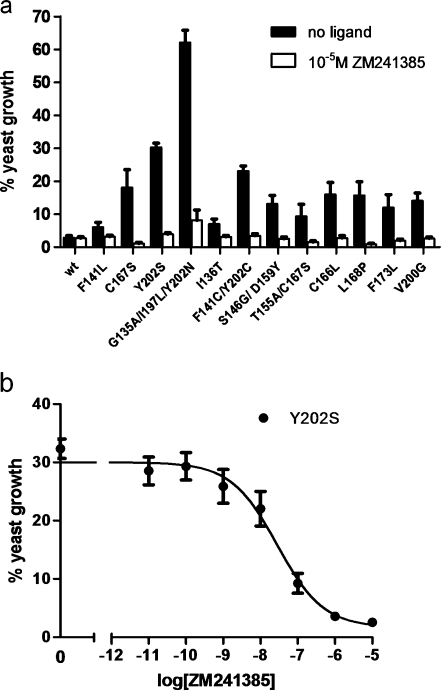
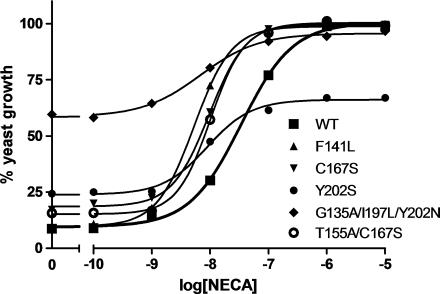
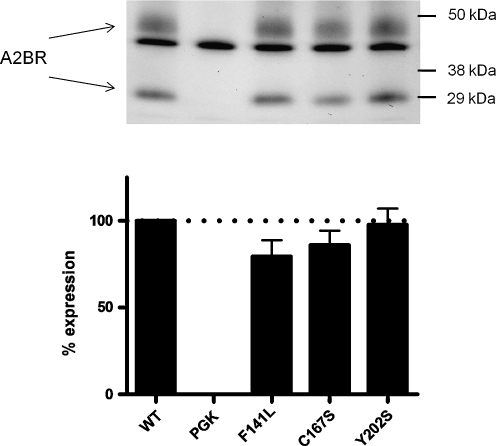
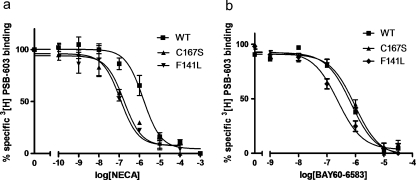
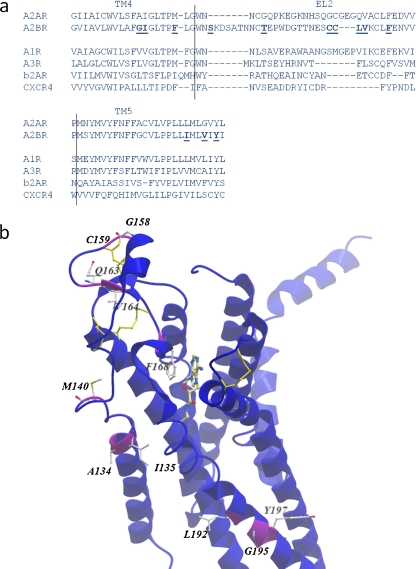
G protein-coupled receptors (GPCRs) are a major drug target and can be activated by a range of stimuli, from photons to proteins. Despite the progress made in the last decade in molecular and structural biology, their exact activation mechanism is still unknown. Here we describe new insights in specific regions essential in adenosine A(2B) receptor activation (A(2B)R), a typical class A GPCR. We applied unbiased random mutagenesis on the middle part of the human adenosine A(2B)R, consisting of transmembrane domains 4 and 5 (TM4 and TM5) linked by extracellular loop 2 (EL2), and subsequently screened in a medium-throughput manner for gain-of-function and constitutively active mutants. For that purpose, we used a genetically engineered yeast strain (Saccharomyces cerevisiae MMY24) with growth as a read-out parameter. From the random mutagenesis screen, 12 different mutant receptors were identified that form three distinct clusters; at the top of TM4, in a cysteine-rich region in EL2, and at the intracellular side of TM5. All mutant receptors show a vast increase in agonist potency and most also displayed a significant increase in constitutive activity. None of these residues are supposedly involved in ligand binding directly. As a consequence, it appears that disrupting the relatively "silent" configuration of the wild-type receptor in each of the three clusters readily causes spontaneous receptor activity.
G 蛋白偶联受体(GPCRs)是一个主要的药物靶点,可以被一系列刺激物激活,从光子到蛋白质。尽管在过去十年中,分子和结构生物学取得了进展,但它们的确切激活机制仍不清楚。在这里,我们描述了在腺苷 A(2B)受体(A(2B)R)激活中关键的特定区域的新见解,A(2B)R 是一种典型的 A 类 GPCR。我们在人类腺苷 A(2B)受体的中间部分(由跨膜域 4 和 5(TM4 和 TM5)通过细胞外环 2(EL2)连接组成)上应用了无偏随机诱变,随后以高通量方式筛选功能获得和组成型激活突变体。为此,我们使用了一种经过基因工程改造的酵母菌株(酿酒酵母 MMY24),以生长作为读出参数。从随机诱变筛选中,鉴定出 12 种不同的突变受体,它们形成三个不同的簇;在 TM4 的顶部、EL2 中的富含半胱氨酸区域以及 TM5 的细胞内侧。所有突变受体都表现出激动剂效力的显著增加,大多数也表现出显著的组成型活性增加。这些残基中没有一个被认为直接参与配体结合。因此,似乎破坏三个簇中野生型受体的相对“沉默”构象很容易导致自发的受体活性。