Institute of Microbiology, University Hospital Center and University of Lausanne, Lausanne, Switzerland.
PLoS Pathog. 2011 Sep;7(9):e1002232. doi: 10.1371/journal.ppat.1002232. Epub 2011 Sep 8.
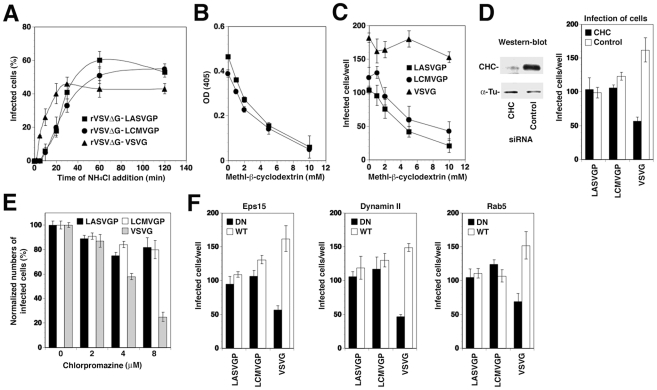
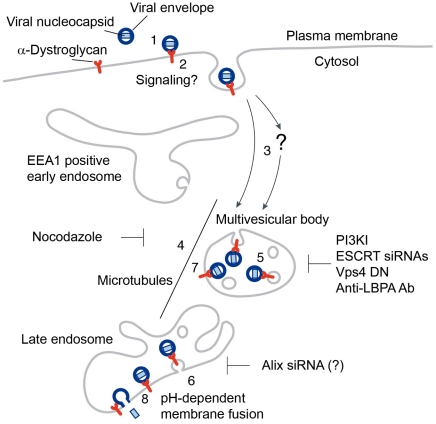
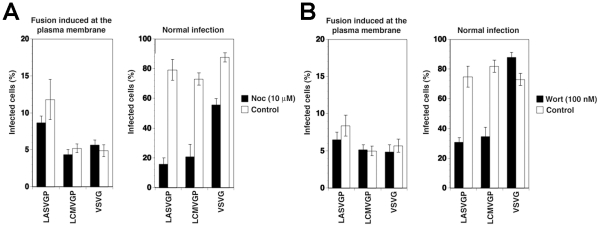
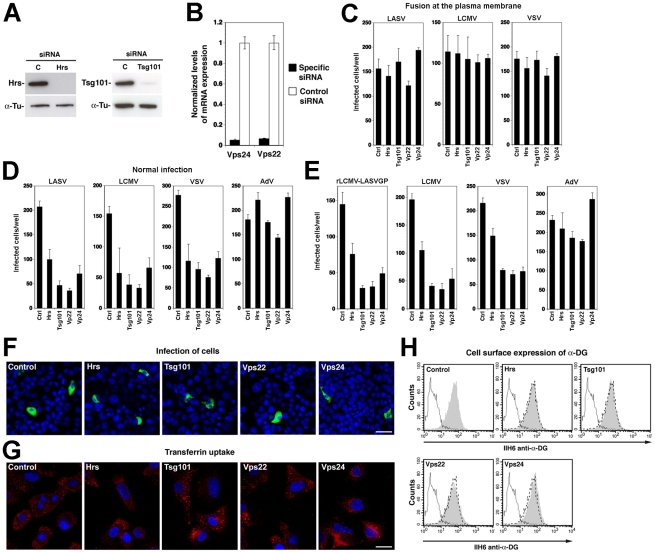
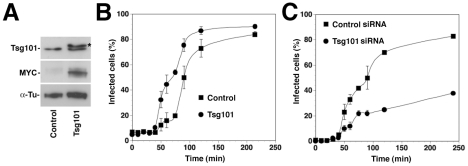
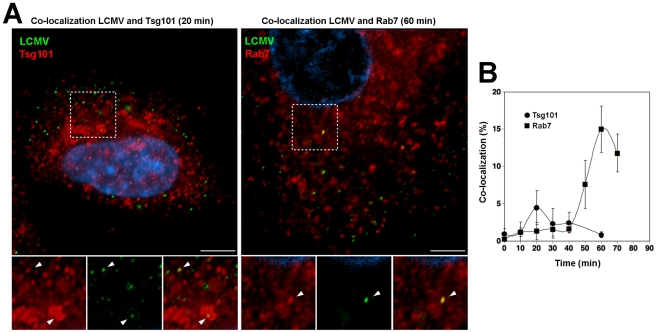
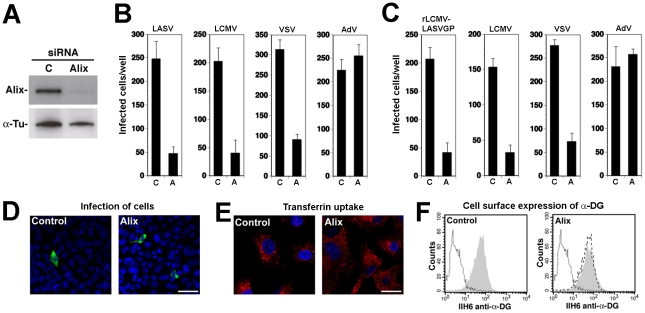
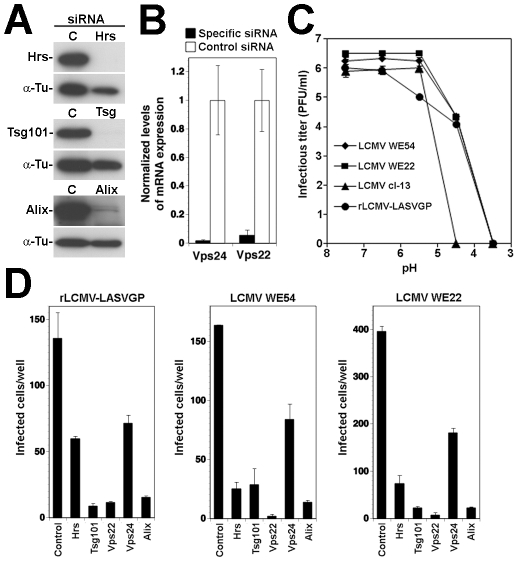
The highly pathogenic Old World arenavirus Lassa virus (LASV) and the prototypic arenavirus lymphocytic choriomeningitis virus (LCMV) use α-dystroglycan as a cellular receptor and enter the host cell by an unusual endocytotic pathway independent of clathrin, caveolin, dynamin, and actin. Upon internalization, the viruses are delivered to acidified endosomes in a Rab5-independent manner bypassing classical routes of incoming vesicular trafficking. Here we sought to identify cellular factors involved in the unusual and largely unknown entry pathway of LASV and LCMV. Cell entry of LASV and LCMV required microtubular transport to late endosomes, consistent with the low fusion pH of the viral envelope glycoproteins. Productive infection with recombinant LCMV expressing LASV envelope glycoprotein (rLCMV-LASVGP) and LCMV depended on phosphatidyl inositol 3-kinase (PI3K) as well as lysobisphosphatidic acid (LBPA), an unusual phospholipid that is involved in the formation of intraluminal vesicles (ILV) of the multivesicular body (MVB) of the late endosome. We provide evidence for a role of the endosomal sorting complex required for transport (ESCRT) in LASV and LCMV cell entry, in particular the ESCRT components Hrs, Tsg101, Vps22, and Vps24, as well as the ESCRT-associated ATPase Vps4 involved in fission of ILV. Productive infection with rLCMV-LASVGP and LCMV also critically depended on the ESCRT-associated protein Alix, which is implicated in membrane dynamics of the MVB/late endosomes. Our study identifies crucial cellular factors implicated in Old World arenavirus cell entry and indicates that LASV and LCMV invade the host cell passing via the MVB/late endosome. Our data further suggest that the virus-receptor complexes undergo sorting into ILV of the MVB mediated by the ESCRT, possibly using a pathway that may be linked to the cellular trafficking and degradation of the cellular receptor.
高致病性旧世界沙粒病毒(LASV)和原型沙粒病毒淋巴细胞性脉络丛脑膜炎病毒(LCMV)使用α- dystroglycan 作为细胞受体,并通过一种不依赖网格蛋白、 caveolin、dynamin 和肌动蛋白的特殊胞吞途径进入宿主细胞。在细胞内吞后,病毒以 Rab5 非依赖性方式被递送到酸化的内体,绕过了传入囊泡运输的经典途径。在这里,我们试图鉴定参与 LASV 和 LCMV 不寻常且大部分未知进入途径的细胞因子。LASV 和 LCMV 的细胞进入需要微管运输到晚期内体,这与病毒包膜糖蛋白的低融合 pH 值一致。用表达 LASV 包膜糖蛋白的重组 LCMV(rLCMV-LASVGP)和 LCMV 进行的有效感染依赖于磷酸肌醇 3-激酶(PI3K)和溶血磷脂酸(LBPA),LBPA 是一种参与晚期内体多泡体(MVB)腔内囊泡(ILV)形成的异常磷脂。我们提供了内体分选复合物所需的运输(ESCRT)在 LASV 和 LCMV 细胞进入中的作用的证据,特别是 ESCRT 成分 Hrs、Tsg101、Vps22 和 Vps24,以及参与 ILV 分裂的 ESCRT 相关 ATP 酶 Vps4。rLCMV-LASVGP 和 LCMV 的有效感染也严重依赖于 ESCRT 相关蛋白 Alix,该蛋白参与 MVB/晚期内体的膜动力学。我们的研究确定了参与旧世界沙粒病毒细胞进入的关键细胞因子,并表明 LASV 和 LCMV 通过 MVB/晚期内体侵入宿主细胞。我们的数据进一步表明,病毒-受体复合物通过 ESCRT 介导的 MVB 中的 ILV 进行分选,可能使用一种与细胞受体的细胞运输和降解相关的途径。