Roy A, Exinger F, Losson R
Laboratoire de Génétique, Institut de Biologie Moléculaire et Cellulaire, Strasbourg, France.
Mol Cell Biol. 1990 Oct;10(10):5257-70. doi: 10.1128/mcb.10.10.5257-5270.1990.
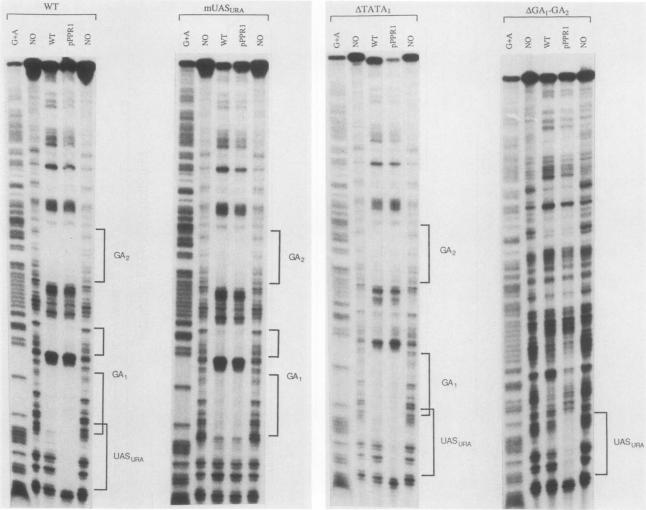
Expression of the yeast pyrimidine biosynthetic gene, URA3, is induced three- to fivefold in response to uracil starvation, and this regulation is mediated by the transcriptional activator PPR1 (pyrimidine pathway regulator 1). In this study, we have analyzed the regulatory elements of the URA3 promoter by DNase I footprinting, using partially purified yeast cell extracts, by deletion mutagenesis, and by 5'-end mapping of RNA transcripts. Two DNA-binding activities have been detected, and at least four distinct cis-acting regions have been identified. A region rich in poly(dA-dT) serves as an upstream promoter element necessary for the basal level of URA3 expression. A 16-base-pair sequence with dyad symmetry acts acts as a uracil-controlled upstream activating site (UASURA) and shows a specific binding only with cell extracts from strains overproducing PPR1. This in vitro binding does not require dihydroorotic acid, the physiological inducer of URA3. The TATA region appears to be composed of two functionally distinct (constitutive and regulatory) elements. Two G + A-rich regions surrounding this TATA box bind an unidentified factor called GA-binding factor. The 5' copy, GA1, is involved in PPR1 induction and overlaps the constitutive TATA region. The 3' region, GA2, is necessary for maximal expression. Neither of these GA sequences acts as a UAS in a CYC1-lacZ context. The promoters of the unlinked but coordinately regulated URA1 and URA4 genes contain highly conserved copies of the UASURA sequence, which prompted us to investigate the effects of many point mutations within this UASURA sequence on PPR1-dependent binding. In this way, we have identified the most important residues of this binding site and found that a nonsymmetrical change of these bases is sufficient to prevent the specific binding and to suppress the UASURA activity in vivo. In addition, we showed that UASURA contains a constitutive activating element which can stimulate transcription from a heterologous promoter independently of dihydroorotic acid and PPR1.
酵母嘧啶生物合成基因URA3的表达在尿嘧啶饥饿时被诱导3至5倍,这种调控由转录激活因子PPR1(嘧啶途径调节因子1)介导。在本研究中,我们通过DNA酶I足迹法,使用部分纯化的酵母细胞提取物,通过缺失诱变以及RNA转录本的5'端图谱分析,对URA3启动子的调控元件进行了分析。检测到两种DNA结合活性,并鉴定出至少四个不同的顺式作用区域。富含聚(dA-dT)的区域作为URA3基础表达水平所必需的上游启动子元件。一个具有二元对称性的16碱基对序列作为尿嘧啶控制的上游激活位点(UASURA),并且仅与过量表达PPR1的菌株的细胞提取物显示特异性结合。这种体外结合不需要二氢乳清酸,即URA3的生理诱导剂。TATA区域似乎由两个功能不同(组成型和调节型)的元件组成。围绕该TATA框的两个富含G + A的区域结合一种未鉴定的因子,称为GA结合因子。5'拷贝GA1参与PPR1诱导并与组成型TATA区域重叠。3'区域GA2是最大表达所必需的。在CYC1-lacZ背景下,这些GA序列均不作为UAS起作用。未连锁但协同调节的URA1和URA4基因的启动子包含UASURA序列的高度保守拷贝,这促使我们研究该UASURA序列内许多点突变对PPR1依赖性结合的影响。通过这种方式,我们鉴定出了该结合位点最重要的残基,并发现这些碱基的非对称变化足以阻止特异性结合并在体内抑制UASURA活性。此外,我们表明UASURA包含一个组成型激活元件,其可以独立于二氢乳清酸和PPR1刺激来自异源启动子的转录。