Department of Microbiology and Molecular Genetics, The University of Texas Health Science Center at Houston, Houston, Texas, United States of America.
PLoS Pathog. 2011 Dec;7(12):e1002453. doi: 10.1371/journal.ppat.1002453. Epub 2011 Dec 22.
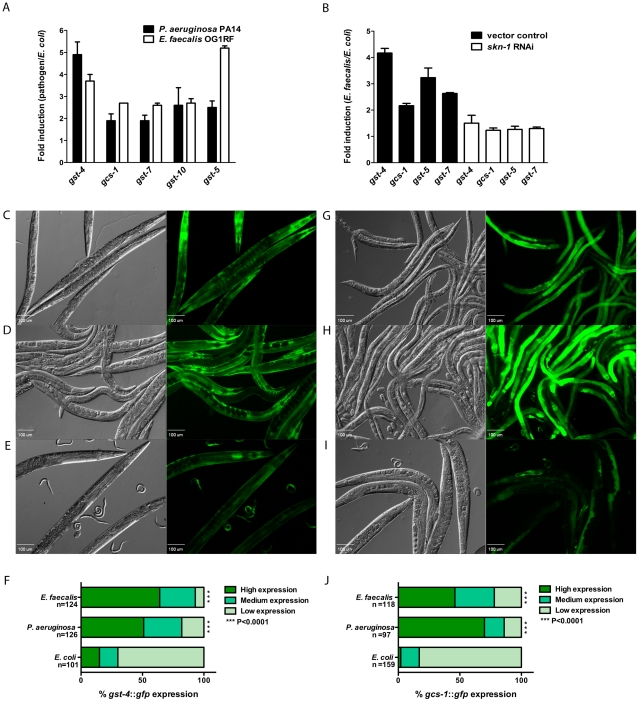
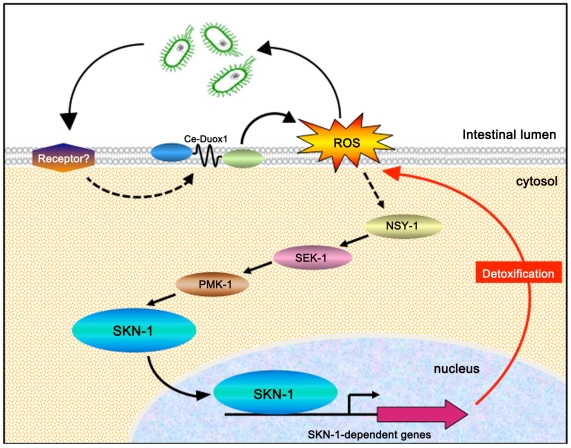
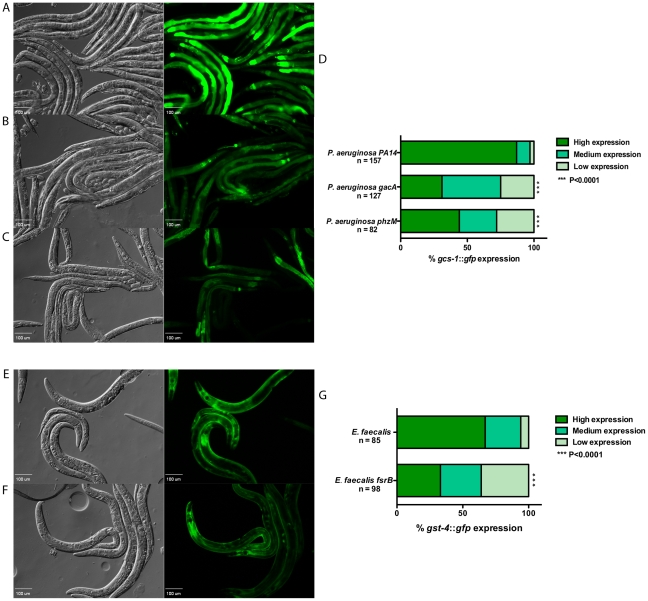
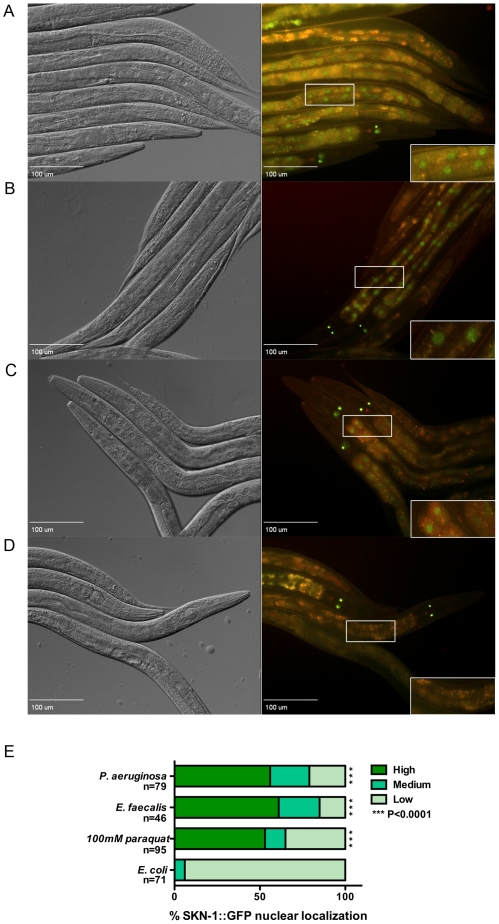
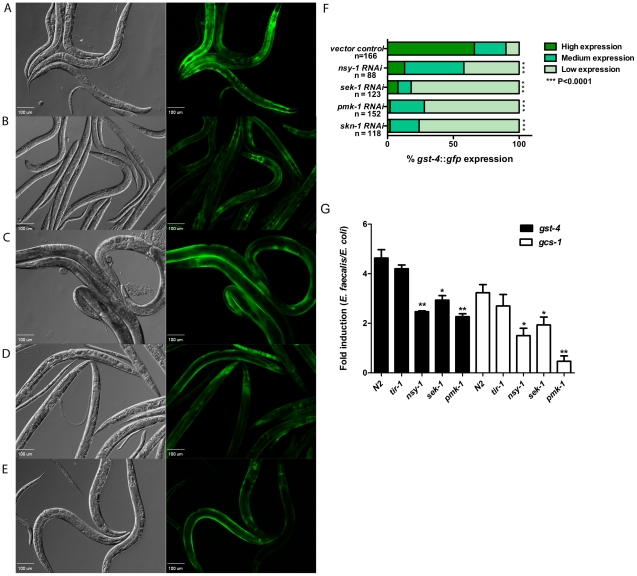
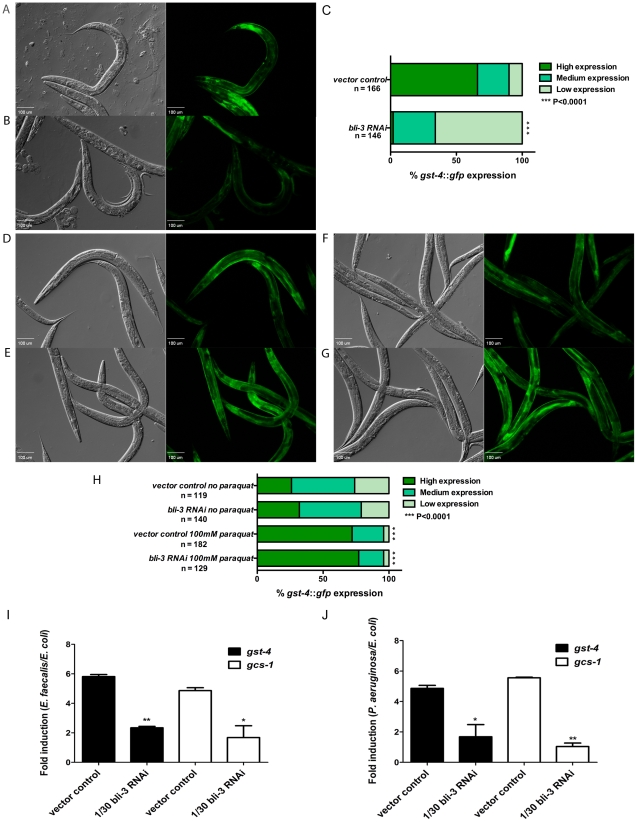
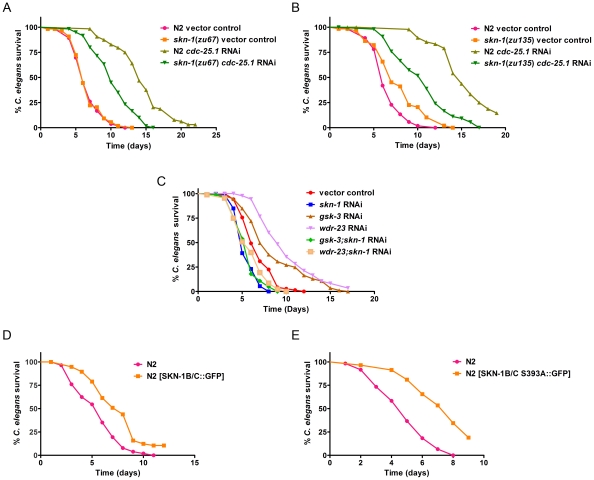
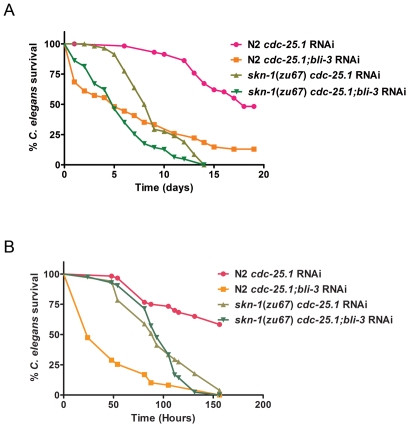
Infected animals will produce reactive oxygen species (ROS) and other inflammatory molecules that help fight pathogens, but can inadvertently damage host tissue. Therefore specific responses, which protect and repair against the collateral damage caused by the immune response, are critical for successfully surviving pathogen attack. We previously demonstrated that ROS are generated during infection in the model host Caenorhabditis elegans by the dual oxidase Ce-Duox1/BLI-3. Herein, an important connection between ROS generation by Ce-Duox1/BLI-3 and upregulation of a protective transcriptional response by SKN-1 is established in the context of infection. SKN-1 is an ortholog of the mammalian Nrf transcription factors and has previously been documented to promote survival, following oxidative stress, by upregulating genes involved in the detoxification of ROS and other reactive compounds. Using qRT-PCR, transcriptional reporter fusions, and a translational fusion, SKN-1 is shown to become highly active in the C. elegans intestine upon exposure to the human bacterial pathogens, Enterococcus faecalis and Pseudomonas aeruginosa. Activation is dependent on the overall pathogenicity of the bacterium, demonstrated by a weakened response observed in attenuated mutants of these pathogens. Previous work demonstrated a role for p38 MAPK signaling both in pathogen resistance and in activating SKN-1 upon exposure to chemically induced oxidative stress. We show that NSY-1, SEK-1 and PMK-1 are also required for SKN-1 activity during infection. Evidence is also presented that the ROS produced by Ce-Duox1/BLI-3 is the source of SKN-1 activation via p38 MAPK signaling during infection. Finally, for the first time, SKN-1 activity is shown to be protective during infection; loss of skn-1 decreases resistance, whereas increasing SKN-1 activity augments resistance to pathogen. Overall, a model is presented in which ROS generation by Ce-Duox1/BLI-3 activates a protective SKN-1 response via p38 MAPK signaling.
感染的动物会产生活性氧(ROS)和其他炎症分子,这些物质有助于抵抗病原体,但也会无意中损害宿主组织。因此,针对免疫反应引起的附带损伤进行保护和修复的特定反应,对于成功抵御病原体攻击至关重要。我们之前证明,在模式宿主秀丽隐杆线虫中,双氧化酶 Ce-Duox1/BLI-3 在感染过程中会产生 ROS。在此,在感染背景下,建立了 ROS 由 Ce-Duox1/BLI-3 产生与 SKN-1 上调保护性转录反应之间的重要联系。SKN-1 是哺乳动物 Nrf 转录因子的同源物,先前已被证明通过上调参与 ROS 和其他反应性化合物解毒的基因,在氧化应激后促进生存。通过 qRT-PCR、转录报告融合和翻译融合,证明 SKN-1 在暴露于人类细菌病原体粪肠球菌和铜绿假单胞菌时,在秀丽隐杆线虫的肠道中高度活跃。激活依赖于细菌的整体致病性,这从这些病原体的减毒突变体中观察到的反应减弱得到证明。先前的工作表明,p38 MAPK 信号通路在病原体抗性以及在暴露于化学诱导的氧化应激时激活 SKN-1 中都起着作用。我们表明,NSY-1、SEK-1 和 PMK-1 在感染期间也需要 SKN-1 活性。还提出了证据表明,Ce-Duox1/BLI-3 产生的 ROS 是通过 p38 MAPK 信号通路在感染过程中激活 SKN-1 的来源。最后,首次证明 SKN-1 活性在感染过程中具有保护作用;skn-1 的缺失会降低抗性,而增加 SKN-1 活性则会增强对病原体的抗性。总的来说,提出了一个模型,其中 Ce-Duox1/BLI-3 产生的 ROS 通过 p38 MAPK 信号通路激活保护性 SKN-1 反应。