Centre for Molecular and Biomolecular Informatics, Nijmegen Centre for Molecular Life Sciences, Radboud University Nijmegen Medical Centre, Nijmegen, The Netherlands.
Mol Biol Evol. 2012 Nov;29(11):3497-512. doi: 10.1093/molbev/mss157. Epub 2012 Jun 11.
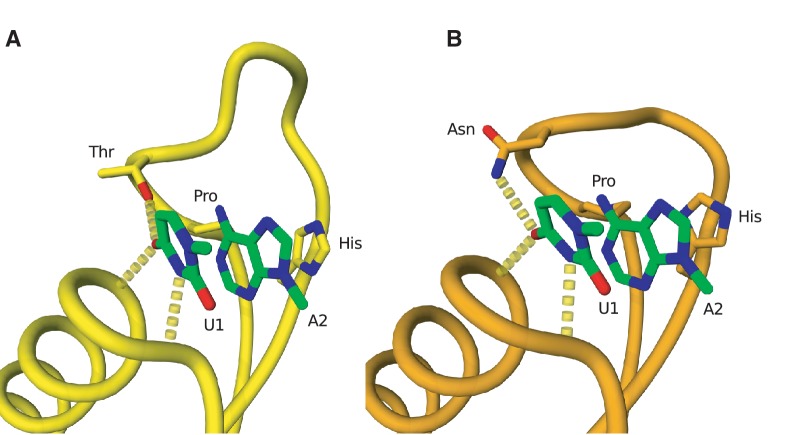
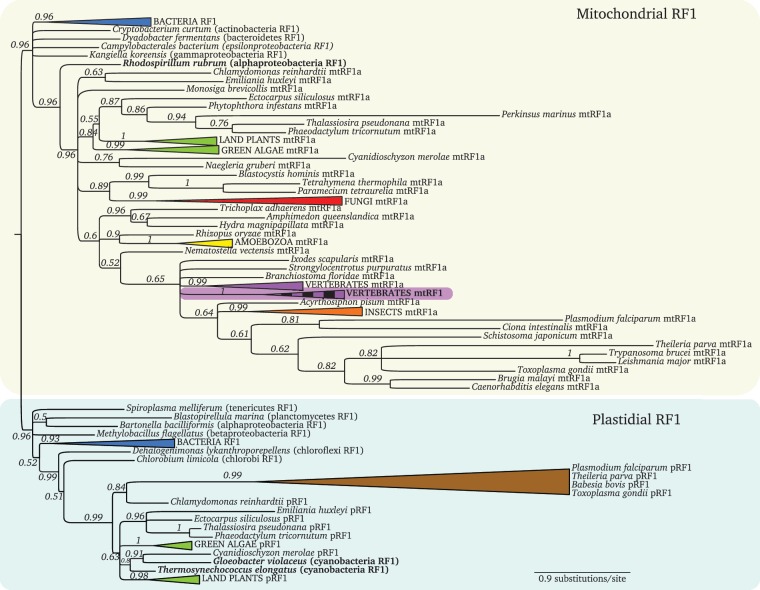
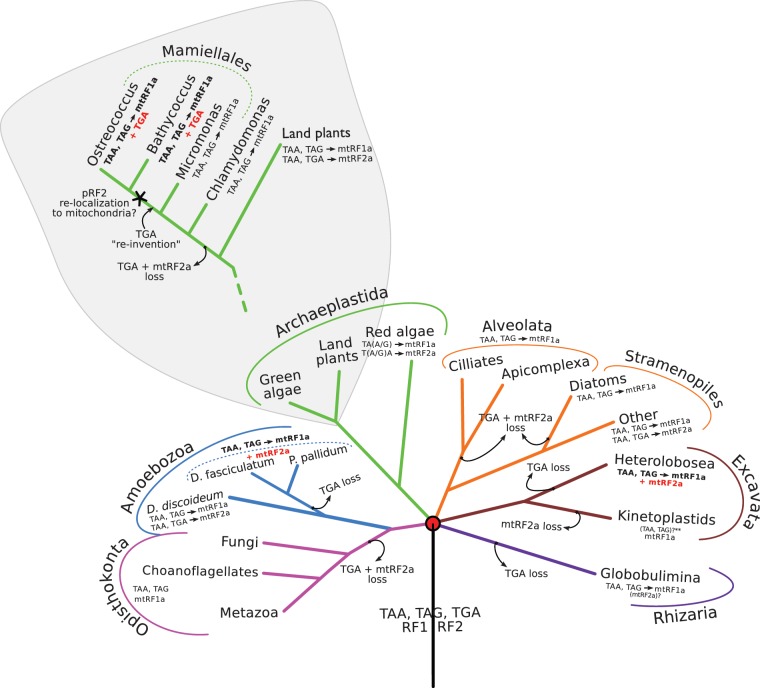
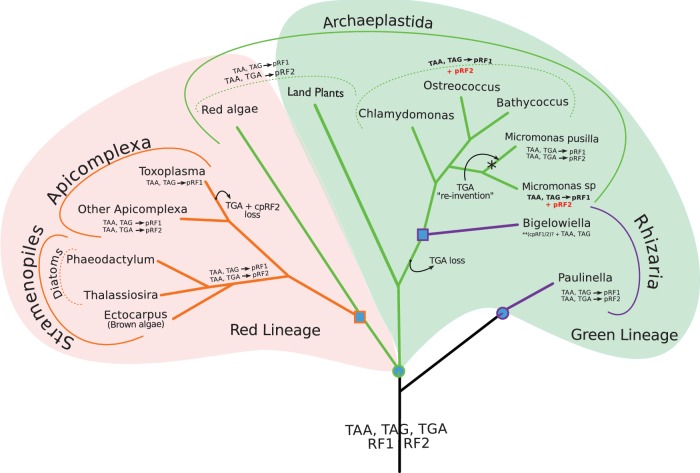
Translation termination is accomplished by proteins of the Class I release factor family (RF) that recognize stop codons and catalyze the ribosomal release of the newly synthesized peptide. Bacteria have two canonical RFs: RF1 recognizes UAA and UAG, RF2 recognizes UAA and UGA. Despite that these two release factor proteins are sufficient for de facto translation termination, the eukaryotic organellar RF protein family, which has evolved from bacterial release factors, has expanded considerably, comprising multiple subfamilies, most of which have not been functionally characterized or formally classified. Here, we integrate multiple sources of information to analyze the remarkable differentiation of the RF family among organelles. We document the origin, phylogenetic distribution and sequence structure features of the mitochondrial and plastidial release factors: mtRF1a, mtRF1, mtRF2a, mtRF2b, mtRF2c, ICT1, C12orf65, pRF1, and pRF2, and review published relevant experimental data. The canonical release factors (mtRF1a, mtRF2a, pRF1, and pRF2) and ICT1 are derived from bacterial ancestors, whereas the others have resulted from gene duplications of another release factor. These new RF family members have all lost one or more specific motifs relevant for bona fide release factor function but are mostly targeted to the same organelle as their ancestor. We also characterize the subset of canonical release factor proteins that bear nonclassical PxT/SPF tripeptide motifs and provide a molecular-model-based rationale for their retained ability to recognize stop codons. Finally, we analyze the coevolution of canonical RFs with the organellar genetic code. Although the RF presence in an organelle and its stop codon usage tend to coevolve, we find three taxa that encode an RF2 without using UGA stop codons, and one reverse scenario, where mamiellales green algae use UGA stop codons in their mitochondria without having a mitochondrial type RF2. For the latter, we put forward a "stop-codon reinvention" hypothesis that involves the retargeting of the plastid release factor to the mitochondrion.
翻译终止是由 I 类释放因子家族(RF)的蛋白质完成的,它们识别终止密码子并催化新合成肽的核糖体释放。细菌有两种规范的 RF:RF1 识别 UAA 和 UAG,RF2 识别 UAA 和 UGA。尽管这两种释放因子蛋白足以实现事实上的翻译终止,但真核细胞器 RF 蛋白家族是从细菌释放因子进化而来的,已经大大扩展,包括多个亚家族,其中大多数尚未进行功能表征或正式分类。在这里,我们整合了多个信息来源来分析细胞器中 RF 家族的显著分化。我们记录了线粒体和质体释放因子的起源、系统发育分布和序列结构特征:mtRF1a、mtRF1、mtRF2a、mtRF2b、mtRF2c、ICT1、C12orf65、pRF1 和 pRF2,并回顾了已发表的相关实验数据。规范的释放因子(mtRF1a、mtRF2a、pRF1 和 pRF2)和 ICT1 源自细菌祖先,而其他的则是另一个释放因子基因重复的结果。这些新的 RF 家族成员都失去了一个或多个与真正的释放因子功能相关的特定基序,但大多数都被靶向到与它们祖先相同的细胞器。我们还描述了具有非经典 PxT/SPF 三肽基序的规范释放因子蛋白亚基,并基于分子模型为它们保留识别终止密码子的能力提供了合理的解释。最后,我们分析了规范 RF 与细胞器遗传密码的共同进化。尽管 RF 在细胞器中的存在及其终止密码子的使用往往是共同进化的,但我们发现有三个分类单元编码 RF2 时不使用 UGA 终止密码子,还有一种相反的情况,即黄藻门绿藻在其线粒体中使用 UGA 终止密码子,但没有线粒体类型的 RF2。对于后者,我们提出了一个“终止密码子再发明”假说,涉及质体释放因子向线粒体的重新靶向。