Department of Human Genetics, Emory University School of Medicine, Atlanta, Georgia, United States of America.
PLoS One. 2012;7(6):e38658. doi: 10.1371/journal.pone.0038658. Epub 2012 Jun 21.
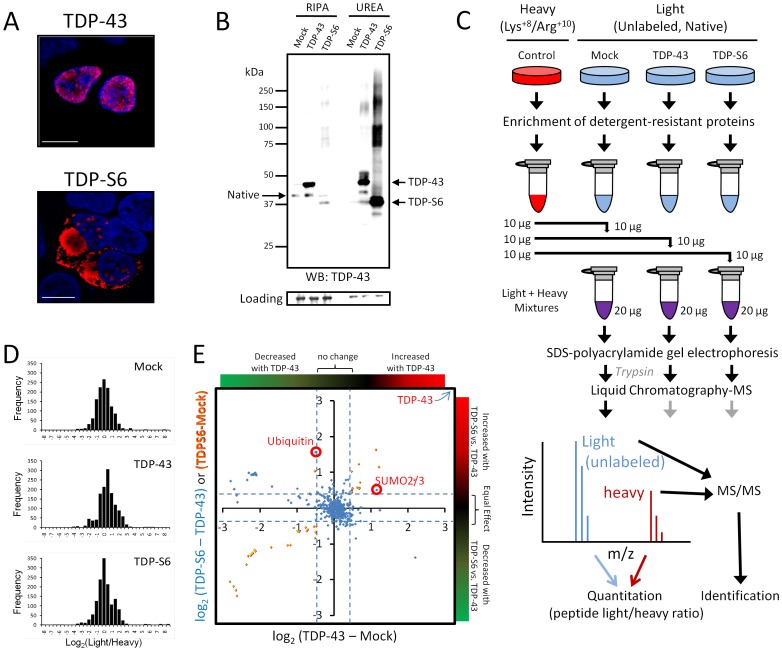
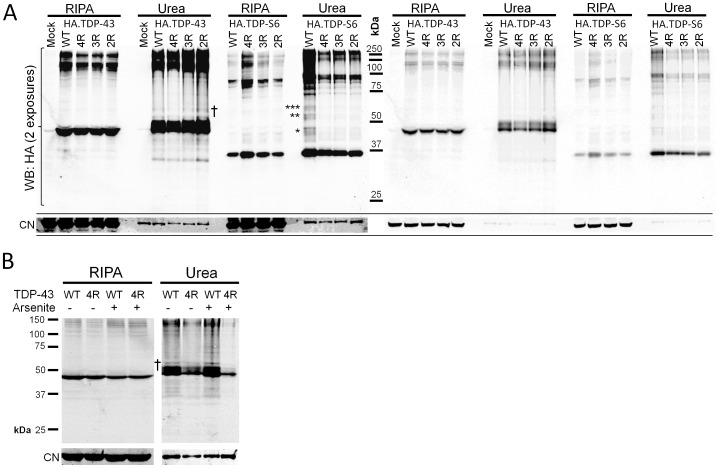
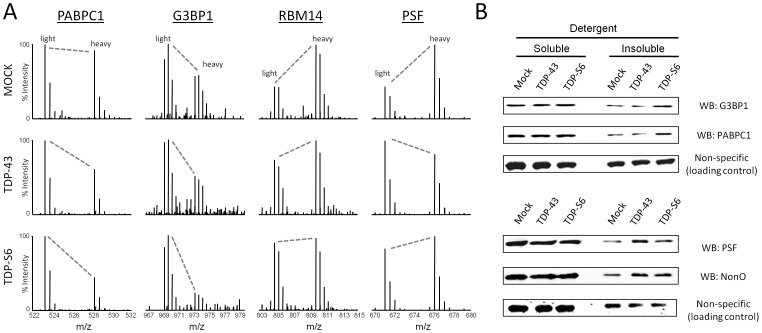
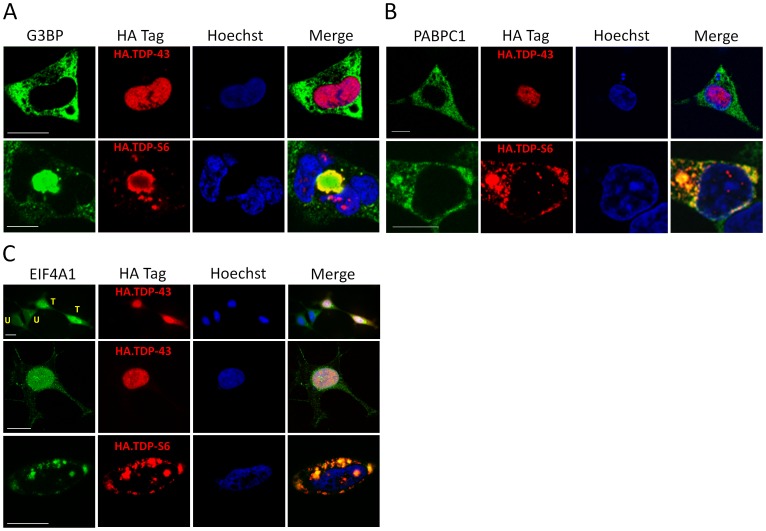
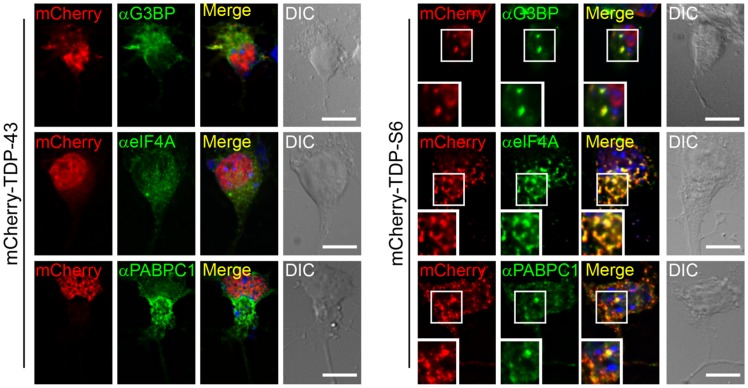
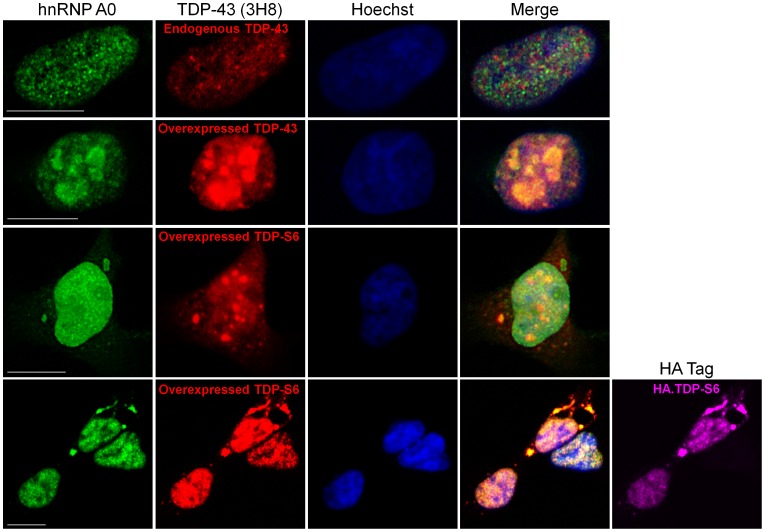
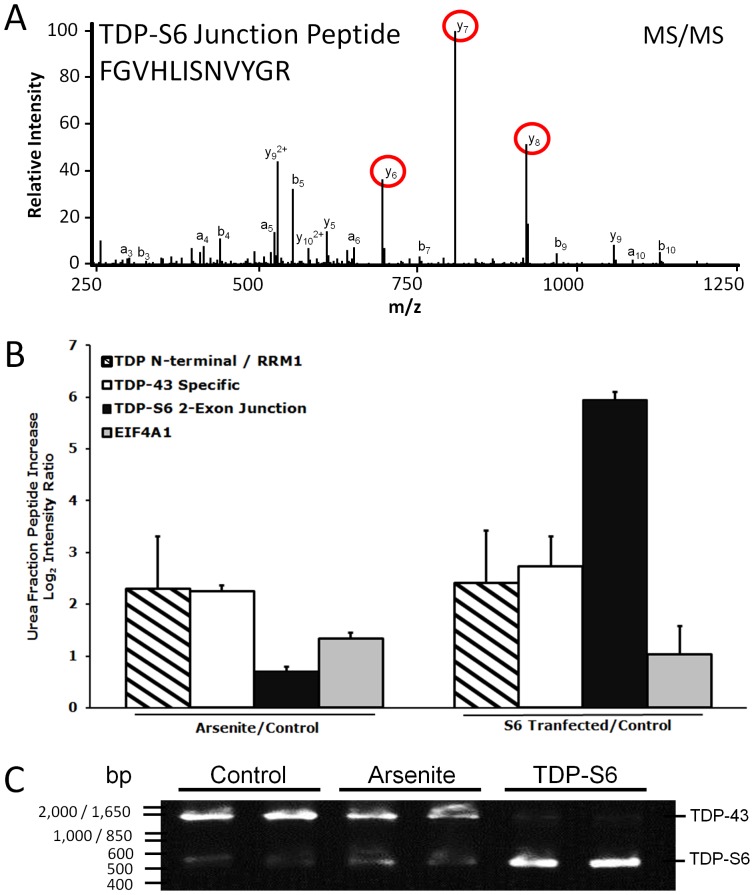
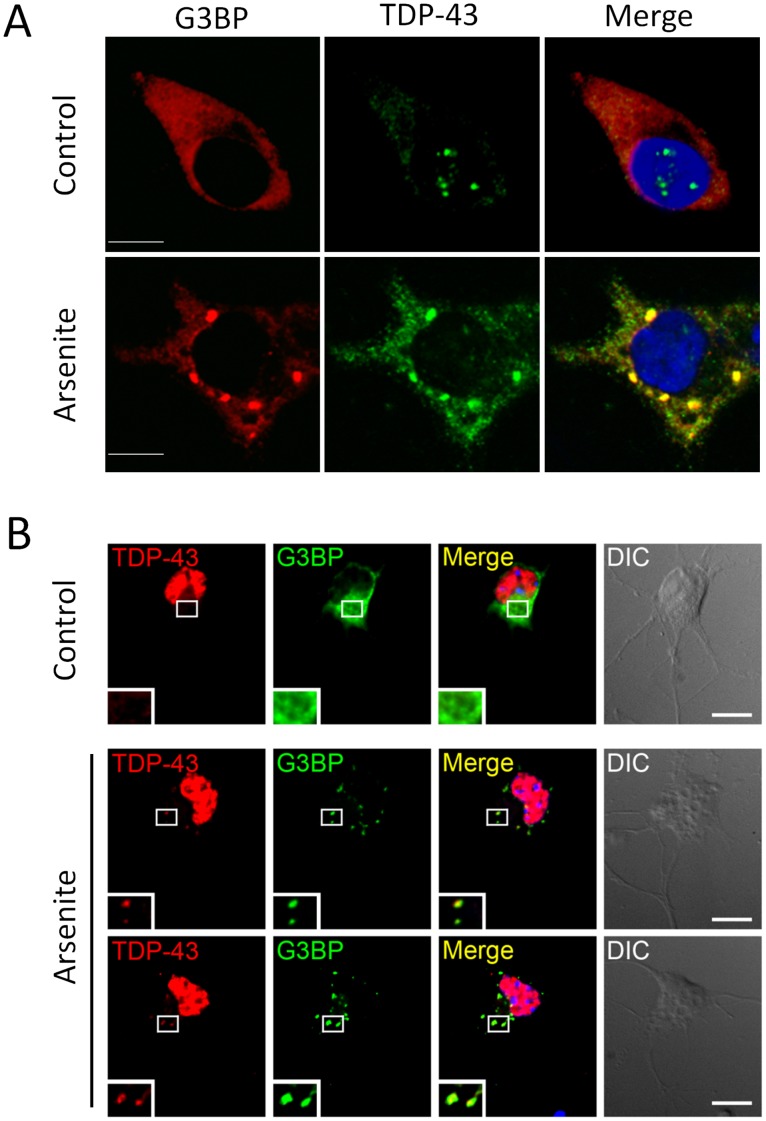
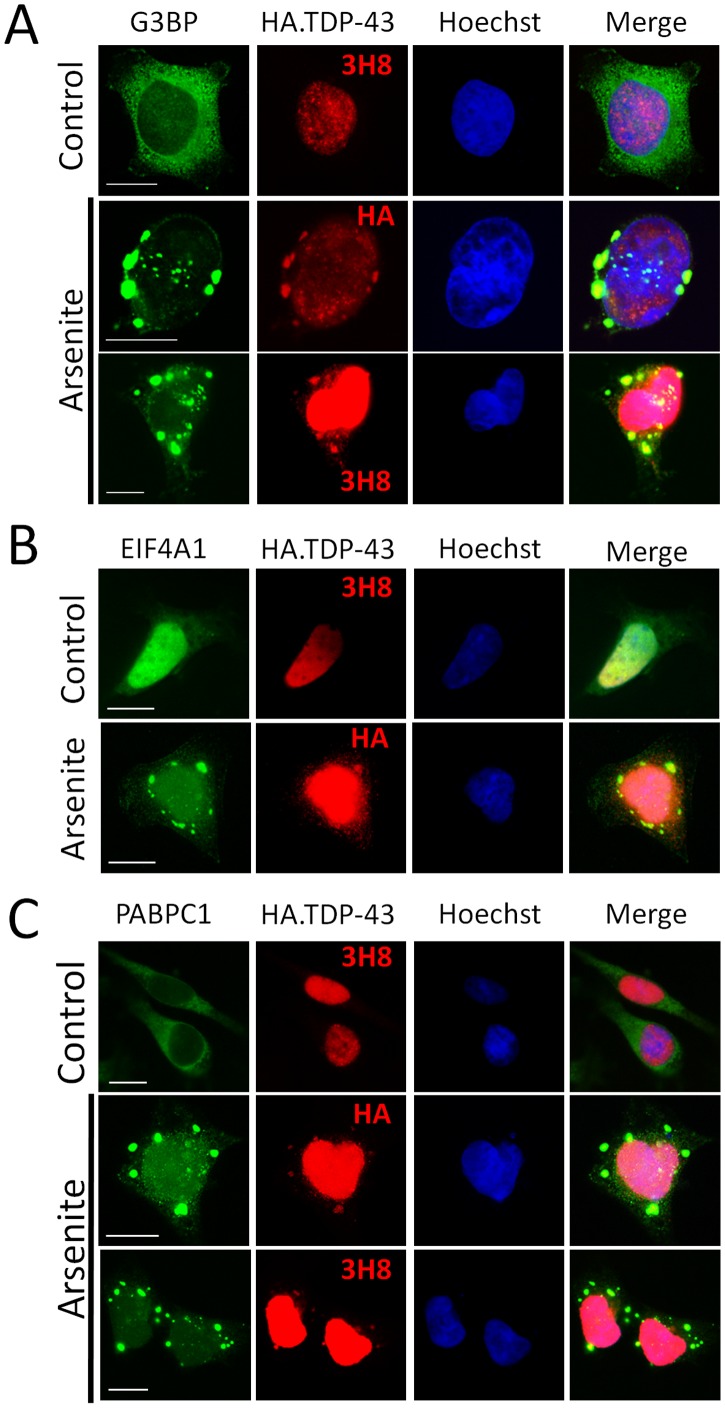
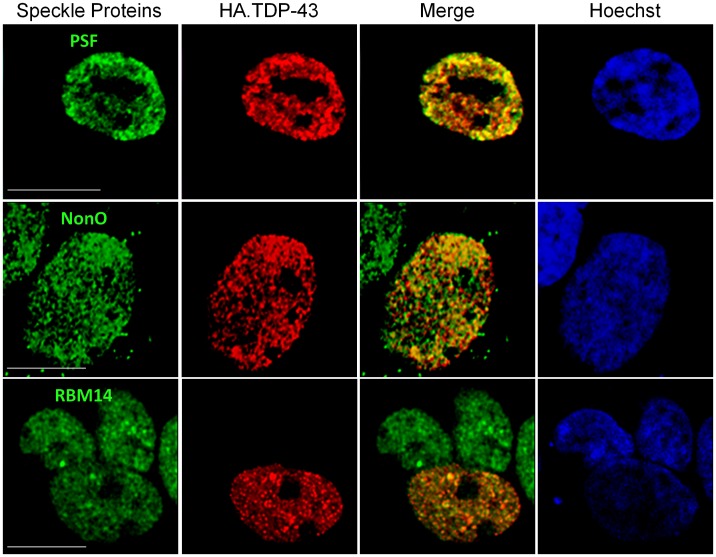
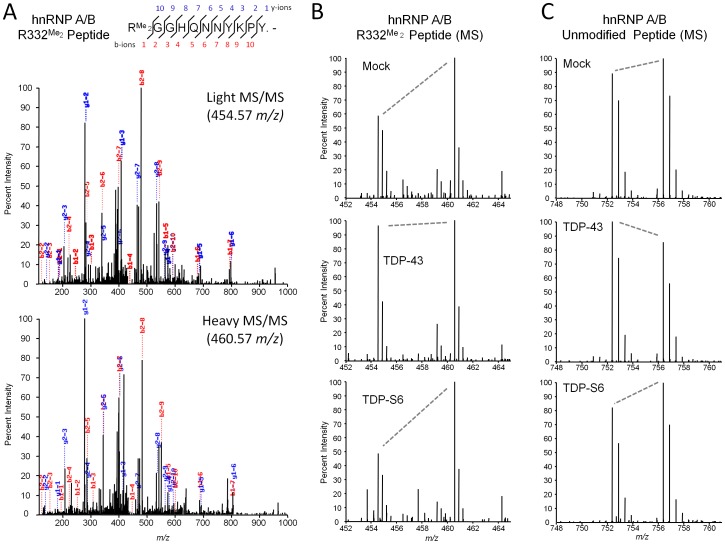
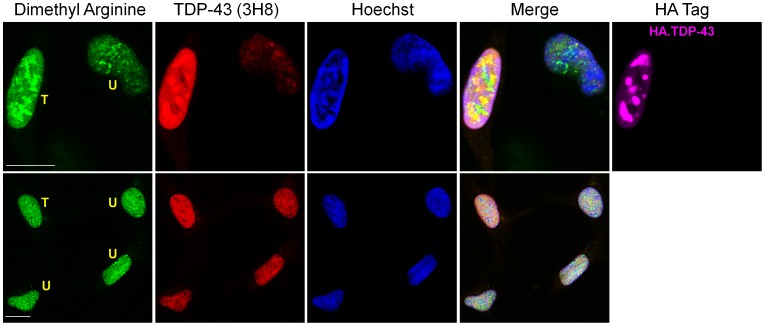
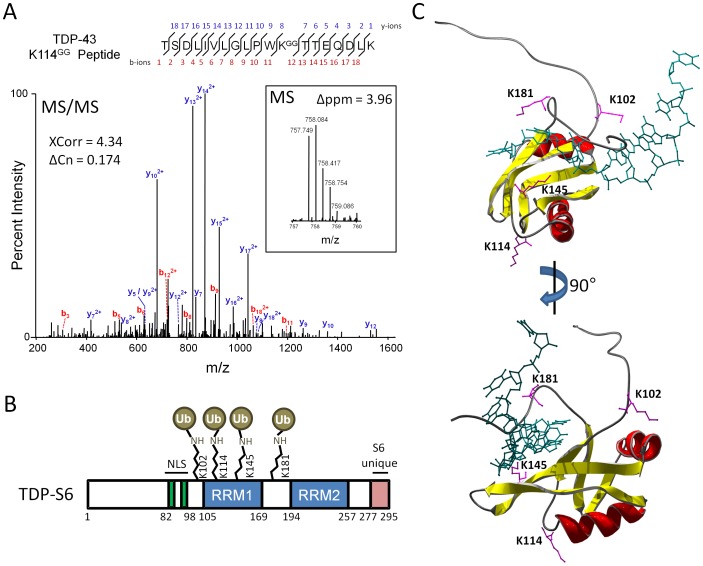
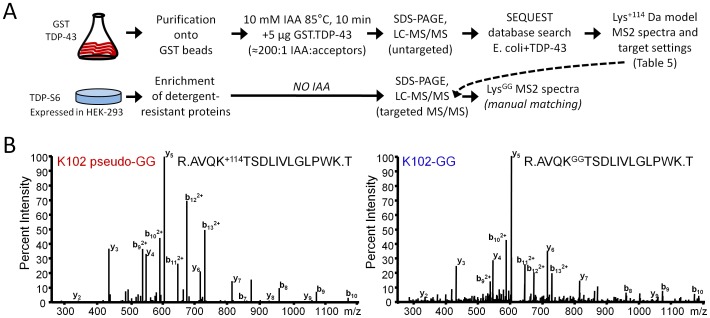
TAR DNA-binding protein 43 (TDP-43) is a major component within ubiquitin-positive inclusions of a number of neurodegenerative diseases that increasingly are considered as TDP-43 proteinopathies. Identities of other inclusion proteins associated with TDP-43 aggregation remain poorly defined. In this study, we identify and quantitate 35 co-aggregating proteins in the detergent-resistant fraction of HEK-293 cells in which TDP-43 or a particularly aggregate prone variant, TDP-S6, were enriched following overexpression, using stable isotope-labeled (SILAC) internal standards and liquid chromatography coupled to tandem mass spectrometry (LC-MS/MS). We also searched for differential post-translational modification (PTM) sites of ubiquitination. Four sites of ubiquitin conjugation to TDP-43 or TDP-S6 were confirmed by dialkylated GST-TDP-43 external reference peptides, occurring on or near RNA binding motif (RRM) 1. RRM-containing proteins co-enriched in cytoplasmic granular structures in HEK-293 cells and primary motor neurons with insoluble TDP-S6, including cytoplasmic stress granule associated proteins G3BP, PABPC1, and eIF4A1. Proteomic evidence for TDP-43 co-aggregation with paraspeckle markers RBM14, PSF and NonO was also validated by western blot and by immunocytochemistry in HEK-293 cells. An increase in peptides from methylated arginine-glycine-glycine (RGG) RNA-binding motifs of FUS/TLS and hnRNPs was found in the detergent-insoluble fraction of TDP-overexpressing cells. Finally, TDP-43 and TDP-S6 detergent-insoluble species were reduced by mutagenesis of the identified ubiquitination sites, even following oxidative or proteolytic stress. Together, these findings define some of the aggregation partners of TDP-43, and suggest that TDP-43 ubiquitination influences TDP-43 oligomerization.
TAR DNA 结合蛋白 43(TDP-43)是许多神经退行性疾病中泛素阳性包涵体的主要成分,这些疾病越来越被认为是 TDP-43 蛋白病。与 TDP-43 聚集相关的其他包涵体蛋白的身份仍未得到很好的定义。在这项研究中,我们使用稳定同位素标记(SILAC)内标和液相色谱-串联质谱(LC-MS/MS),鉴定并定量了 HEK-293 细胞中去污剂不溶部分中与 TDP-43 或特别易于聚集的变体 TDP-S6 共聚集的 35 种蛋白质。我们还搜索了泛素化的差异翻译后修饰(PTM)位点。通过二烷基化 GST-TDP-43 外参肽,证实了 TDP-43 或 TDP-S6 上或附近的 RNA 结合基序(RRM)1 上的 4 个泛素化位点。在 HEK-293 细胞和含有不溶性 TDP-S6 的原代运动神经元中,富含细胞质颗粒结构的 RRM 蛋白,包括细胞质应激颗粒相关蛋白 G3BP、PABPC1 和 eIF4A1。通过 Western blot 和 HEK-293 细胞的免疫细胞化学,也验证了 TDP-43 与核小核糖核蛋白(PAR)标记物 RBM14、PSF 和 NonO 共聚集的蛋白质组学证据。在 TDP 过表达细胞的去污剂不溶部分中,还发现 FUS/TLS 和 hnRNPs 的甲基化精氨酸-甘氨酸-甘氨酸(RGG)RNA 结合基序的肽增加。最后,通过鉴定的泛素化位点的突变,即使在氧化或蛋白水解应激后,TDP-43 和 TDP-S6 去污剂不溶物种的水平也降低了。总之,这些发现定义了 TDP-43 的一些聚集伙伴,并表明 TDP-43 的泛素化影响 TDP-43 的寡聚化。