School of Life Sciences, University of Warwick, Coventry, United Kingdom.
PLoS One. 2012;7(7):e41119. doi: 10.1371/journal.pone.0041119. Epub 2012 Jul 19.
Escherichia coli Shiga-like toxin 1 normally traffics to the endoplasmic reticulum (ER) in sensitive mammalian cells from where the catalytic A chain (SLTxA1) dislocates to the cytosol to inactivate ribosomes. Currently, no molecular details of the dislocation process are available. To investigate the mechanism of the dislocation step we expressed SLTxA1 in the ER of Saccharomyces cerevisiae.
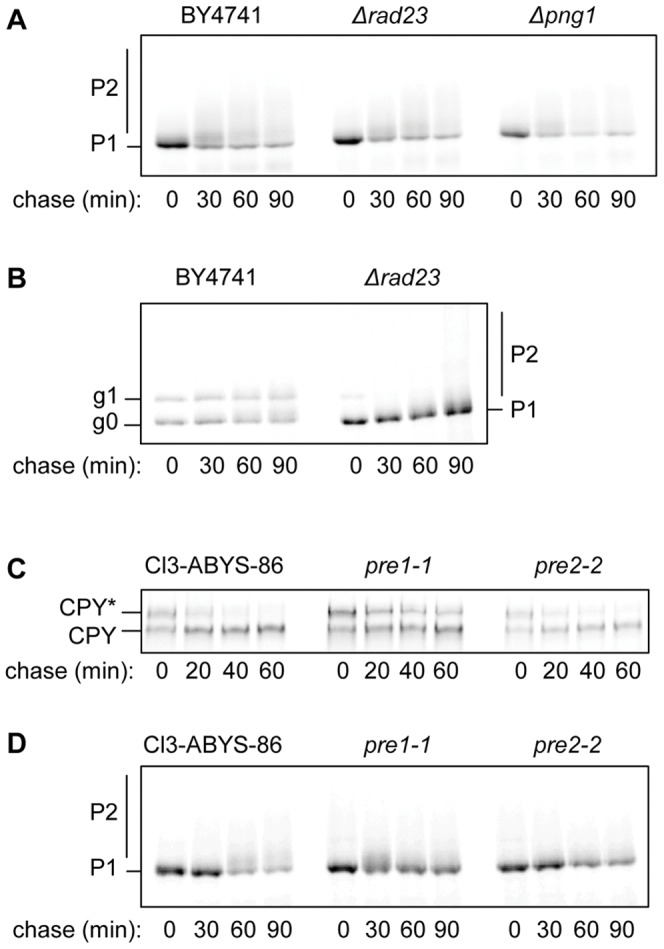
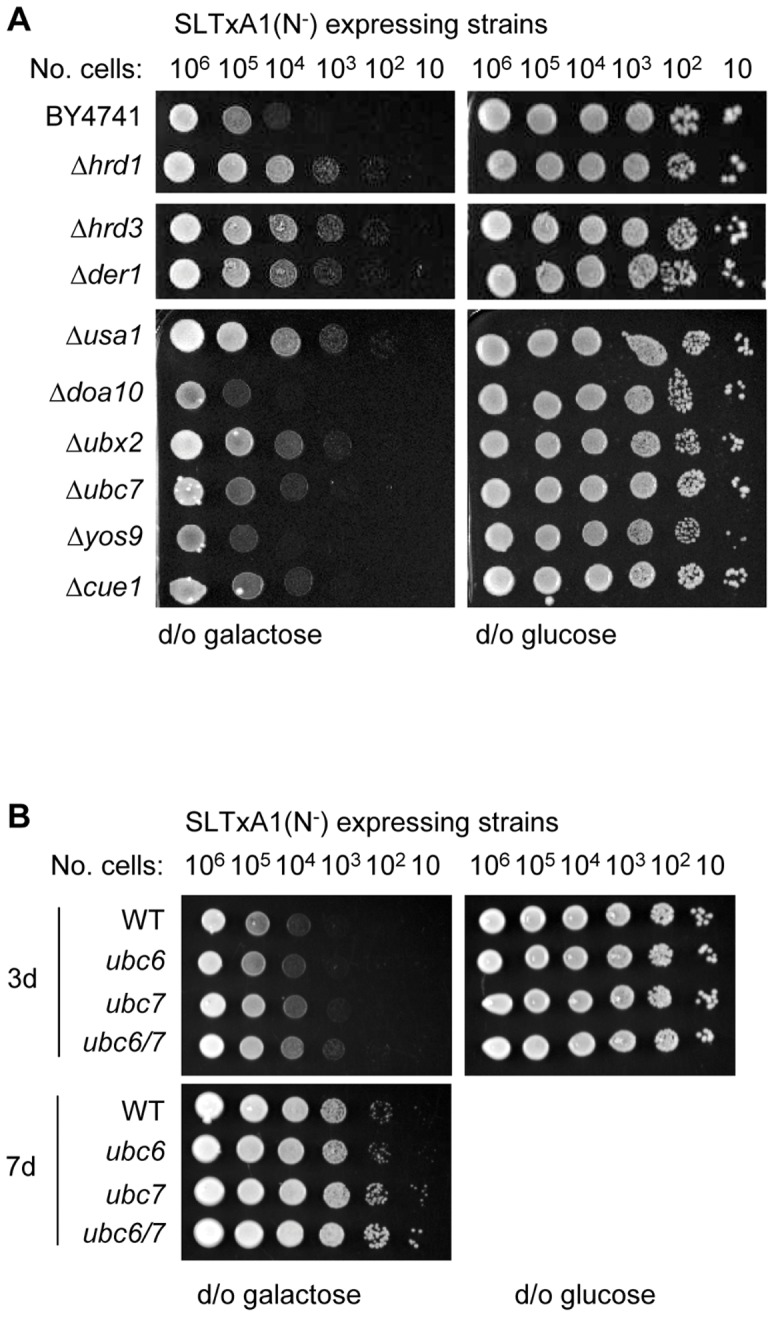
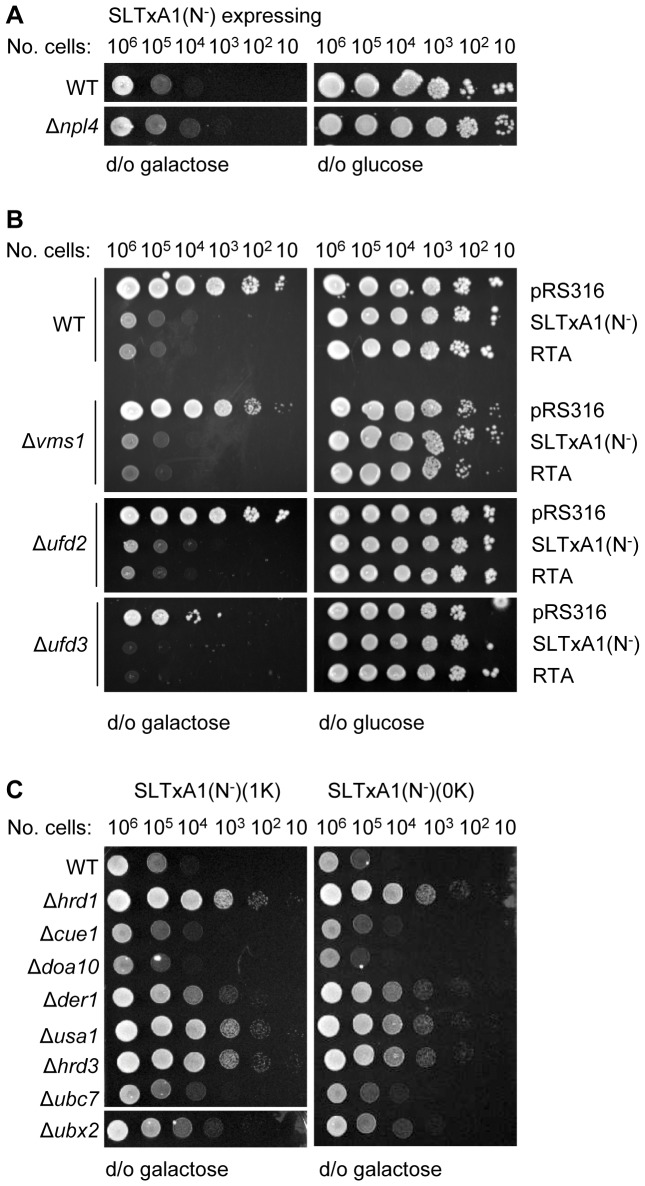
Using a combination of growth studies and biochemical tracking in yeast knock-out strains we show that SLTxA1 follows an ER-associated degradation (ERAD) pathway to enter the cytosol in a step mediated by the transmembrane Hrd1p ubiquitin ligase complex. ER-to-cytosol dislocation of the bulk population of SLTxA1 requires Cdc48p and its ubiquitin-handling co-factor Npl4p, and this population of toxin is terminally dispatched by proteasomal degradation. A small sub-population of SLTxA1 uncouples from this classical ERAD pathway and recovers catalytic activity in the cytosol. The pathway that leads to toxicity is also Hrd1p-dependent but, unlike that for the related ricin A chain toxin, SLTxA1 dislocation does require the catalytic cysteine of Hrd1p. However it does not depend on canonical ubiquitylation since toxin variants lacking endogenous lysyl residues also utilize this pathway, and furthermore there is no requirement for a number of Cdc48p co-factors.
The fraction of SLTxA1 that disengages from the ERAD pathway thus does so upstream of Cdc48p interactions and downstream of Hrd1p interactions, in a step that possibly involves de-ubiquitylation. Mechanistically therefore, the dislocation of this toxin is quite distinct from that of conventional ERAD substrates that are normally degraded, and the toxins partially characterised to date that do not require the catalytic cysteine of the major Hrd1p component of the dislocation apparatus.
大肠杆菌志贺样毒素 1 通常在敏感的哺乳动物细胞中经内质网 (ER) 运输,在那里催化 A 链(SLTxA1)易位到细胞质以失活核糖体。目前,尚无易位过程的分子细节。为了研究易位步骤的机制,我们在酿酒酵母的 ER 中表达了 SLTxA1。
使用生长研究和酵母敲除株的生化追踪相结合,我们表明 SLTxA1 通过内质网相关降解 (ERAD) 途径进入细胞质,这一途径由跨膜 Hrd1p 泛素连接酶复合物介导。大量 SLTxA1 的 ER 到细胞质的易位需要 Cdc48p 和其泛素处理辅助因子 Npl4p,并且该毒素的大部分通过蛋白酶体降解被终末处理。一小部分 SLTxA1 与这种经典的 ERAD 途径解耦,并在细胞质中恢复催化活性。导致毒性的途径也依赖于 Hrd1p,但与相关的蓖麻毒素 A 链毒素不同,SLTxA1 的易位不需要 Hrd1p 的催化半胱氨酸。然而,它不依赖于典型的泛素化,因为缺乏内源性赖氨酸残基的毒素变体也利用这种途径,此外,不需要许多 Cdc48p 辅助因子。
因此,与通常被降解的常规 ERAD 底物相比,从 ERAD 途径脱离的 SLTxA1 部分可能涉及去泛素化,从而从 ERAD 途径脱离的 SLTxA1 部分在 Cdc48p 相互作用之前和 Hrd1p 相互作用之后的步骤中脱离,从机制上讲,这种毒素的易位与目前部分表征的不需要易位装置主要 Hrd1p 成分的催化半胱氨酸的其它毒素的易位明显不同。