Department of Plant Sciences, University of Cambridge, Cambridge, United Kingdom.
PLoS Genet. 2012;8(8):e1002844. doi: 10.1371/journal.pgen.1002844. Epub 2012 Aug 2.
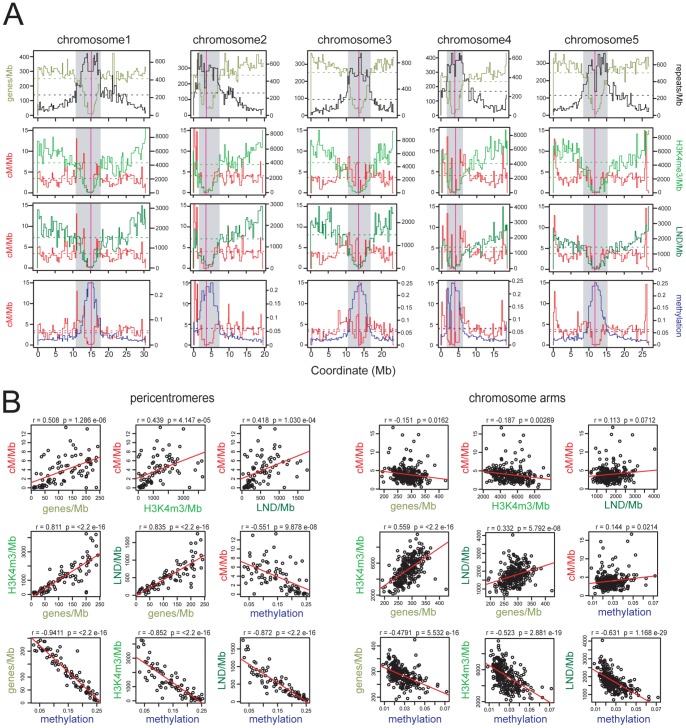
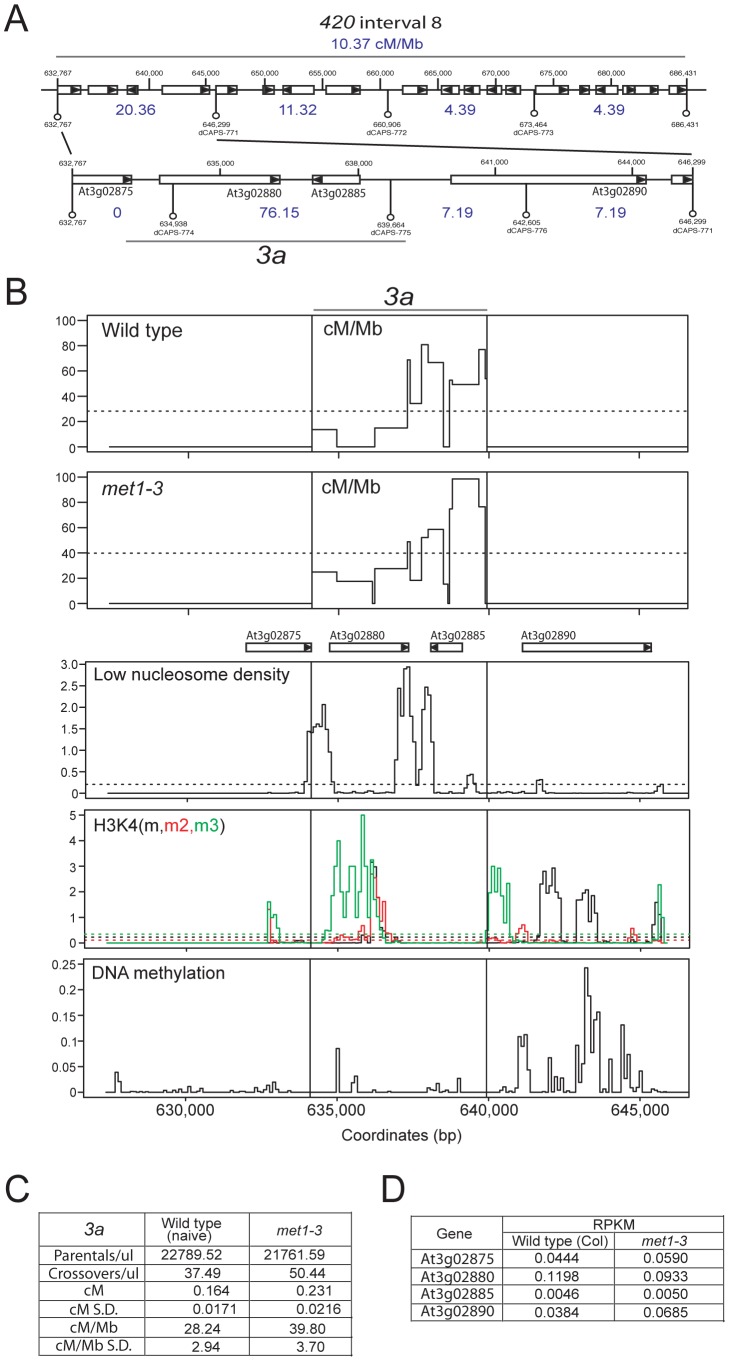
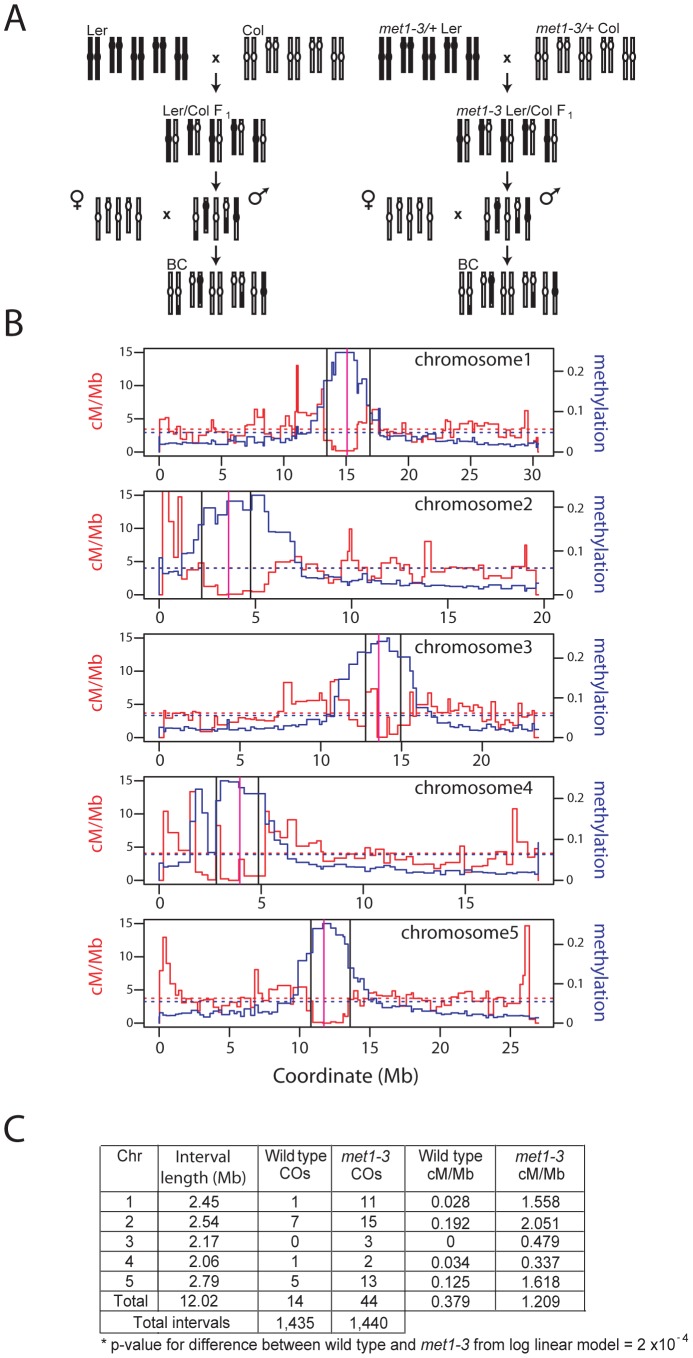
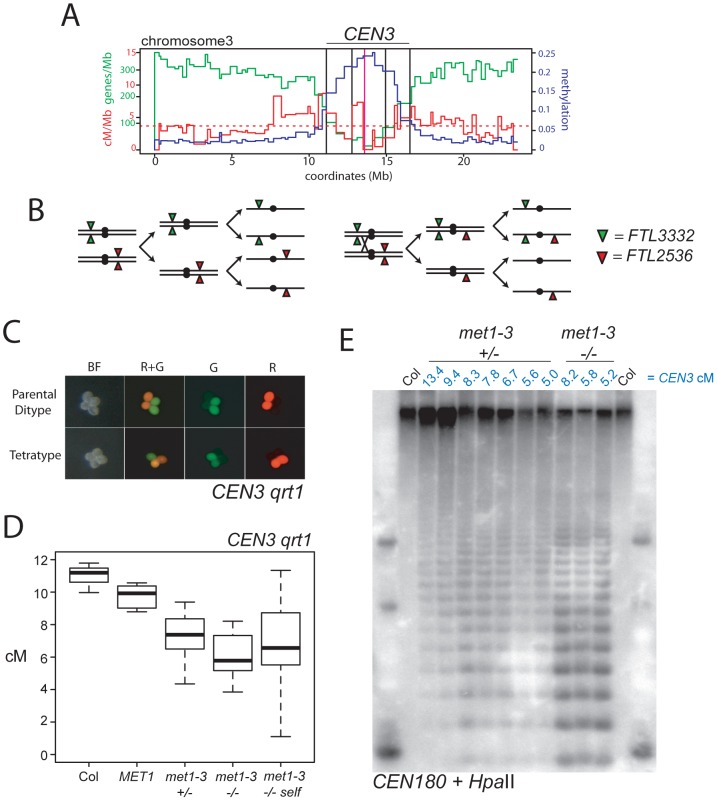
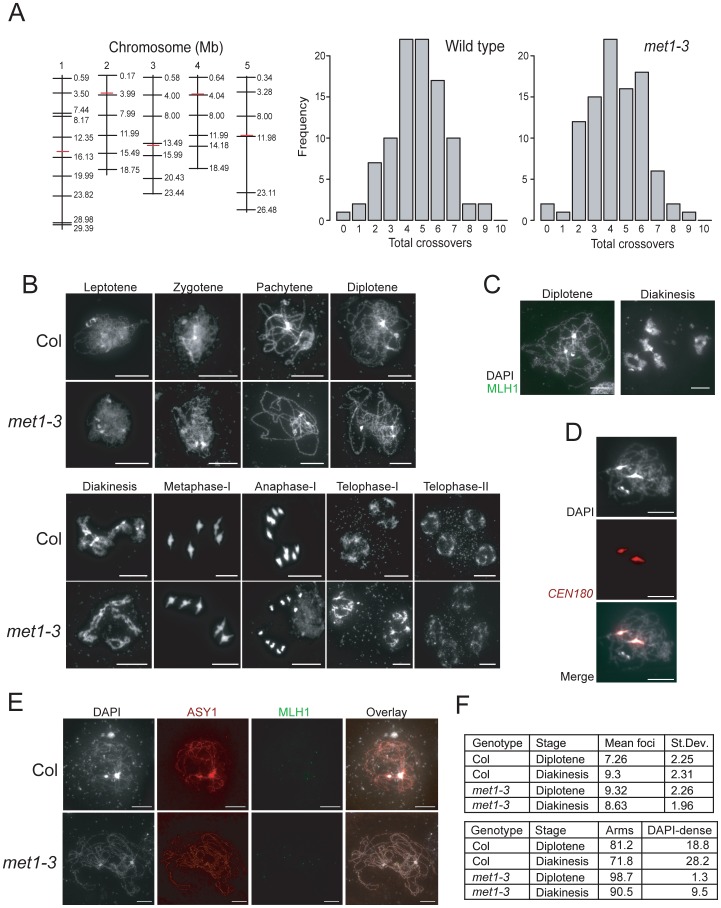
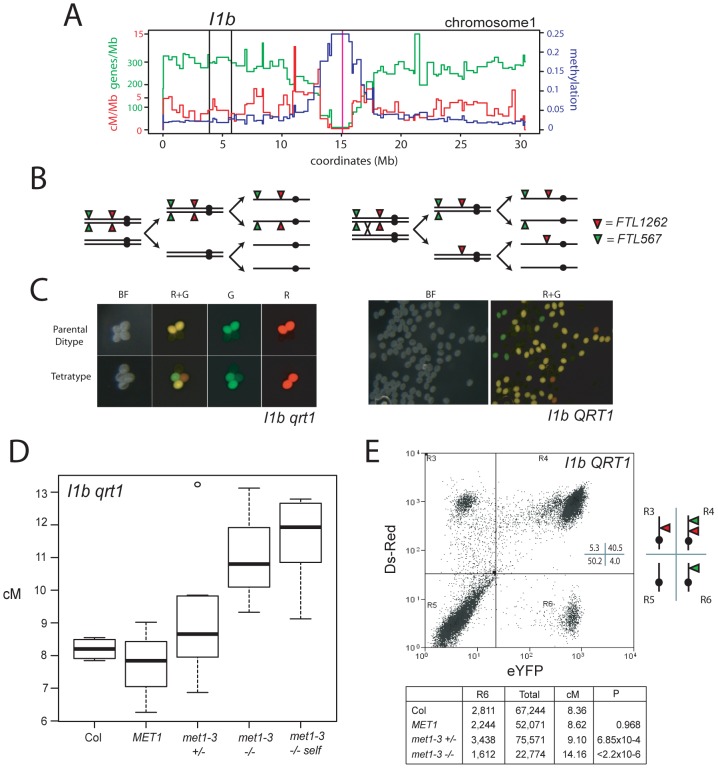
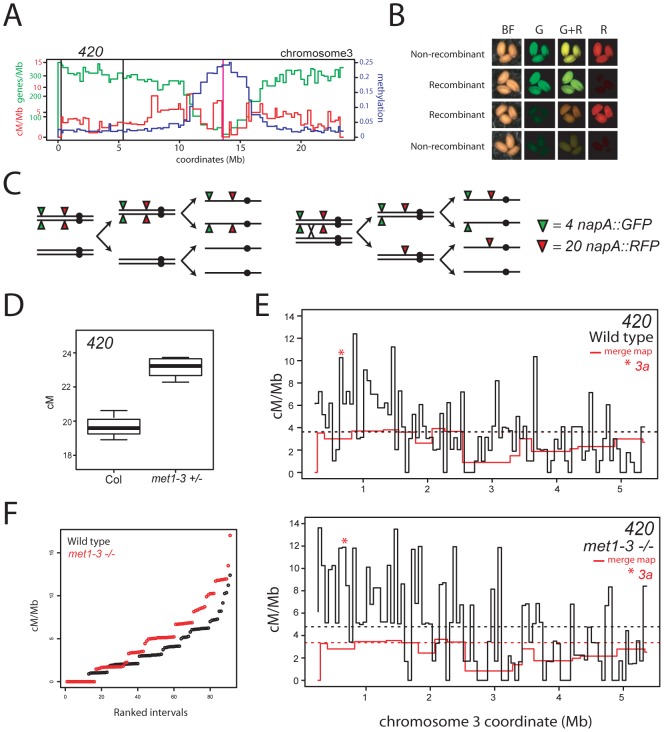
Meiosis is a specialized eukaryotic cell division that generates haploid gametes required for sexual reproduction. During meiosis, homologous chromosomes pair and undergo reciprocal genetic exchange, termed crossover (CO). Meiotic CO frequency varies along the physical length of chromosomes and is determined by hierarchical mechanisms, including epigenetic organization, for example methylation of the DNA and histones. Here we investigate the role of DNA methylation in determining patterns of CO frequency along Arabidopsis thaliana chromosomes. In A. thaliana the pericentromeric regions are repetitive, densely DNA methylated, and suppressed for both RNA polymerase-II transcription and CO frequency. DNA hypomethylated methyltransferase1 (met1) mutants show transcriptional reactivation of repetitive sequences in the pericentromeres, which we demonstrate is coupled to extensive remodeling of CO frequency. We observe elevated centromere-proximal COs in met1, coincident with pericentromeric decreases and distal increases. Importantly, total numbers of CO events are similar between wild type and met1, suggesting a role for interference and homeostasis in CO remodeling. To understand recombination distributions at a finer scale we generated CO frequency maps close to the telomere of chromosome 3 in wild type and demonstrate an elevated recombination topology in met1. Using a pollen-typing strategy we have identified an intergenic nucleosome-free CO hotspot 3a, and we demonstrate that it undergoes increased recombination activity in met1. We hypothesize that modulation of 3a activity is caused by CO remodeling driven by elevated centromeric COs. These data demonstrate how regional epigenetic organization can pattern recombination frequency along eukaryotic chromosomes.
减数分裂是一种专门的真核细胞分裂,它产生的单倍体配子是有性繁殖所必需的。在减数分裂过程中,同源染色体配对并发生相互遗传交换,称为交叉(CO)。减数分裂 CO 频率沿染色体的物理长度而变化,由分层机制决定,包括表观遗传组织,例如 DNA 和组蛋白的甲基化。在这里,我们研究了 DNA 甲基化在决定拟南芥染色体 CO 频率模式中的作用。在拟南芥中,着丝粒周围区域是重复的,高度 DNA 甲基化,并且抑制 RNA 聚合酶-II 转录和 CO 频率。DNA 低甲基化甲基转移酶 1(met1)突变体显示出着丝粒周围重复序列的转录重新激活,我们证明这与 CO 频率的广泛重塑有关。我们观察到 met1 中着丝粒近端 CO 的增加,与着丝粒周围减少和远端增加相一致。重要的是,野生型和 met1 之间的 CO 事件总数相似,这表明干涉和内稳态在 CO 重塑中起作用。为了更精细地了解重组分布,我们在野生型和 met1 中生成了接近染色体 3 端的 CO 频率图谱,并证明了 met1 中重组拓扑的升高。使用花粉分型策略,我们鉴定了一个基因间无核小体 CO 热点 3a,并证明它在 met1 中经历了增加的重组活性。我们假设 3a 活性的调节是由升高的着丝粒 CO 驱动的 CO 重塑引起的。这些数据表明,区域表观遗传组织如何沿真核染色体模式化重组频率。