Gene Regulation and Chromosome Biology Laboratory, Frederick National Laboratory for Cancer Research, Frederick MD, USA.
Front Oncol. 2012 Nov 16;2:166. doi: 10.3389/fonc.2012.00166. eCollection 2012.
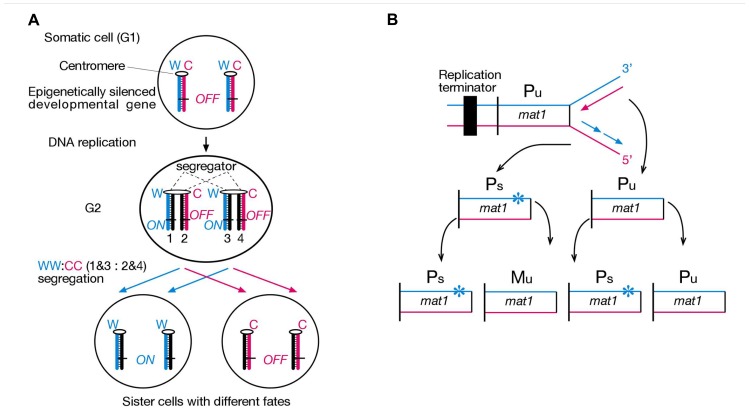
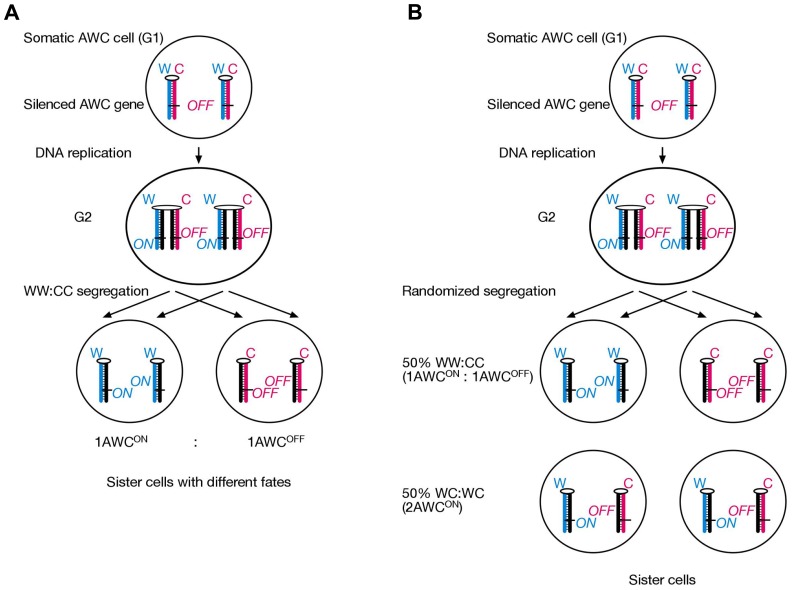
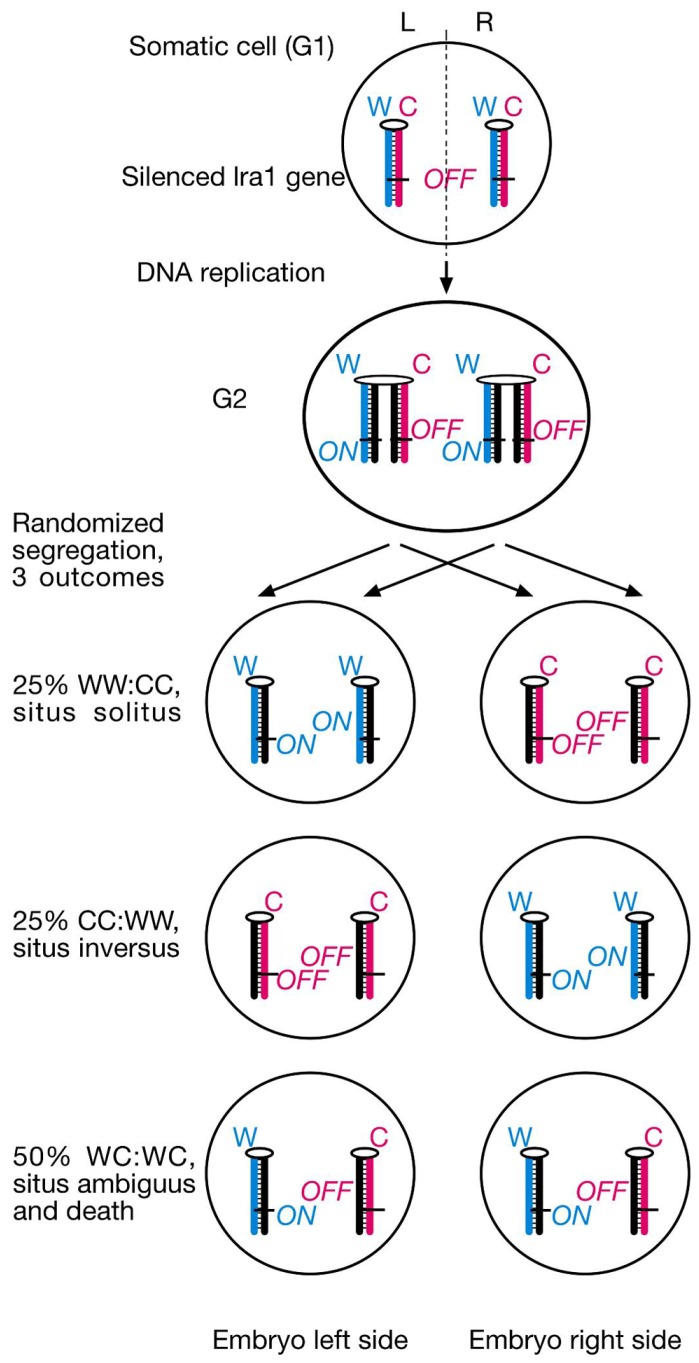
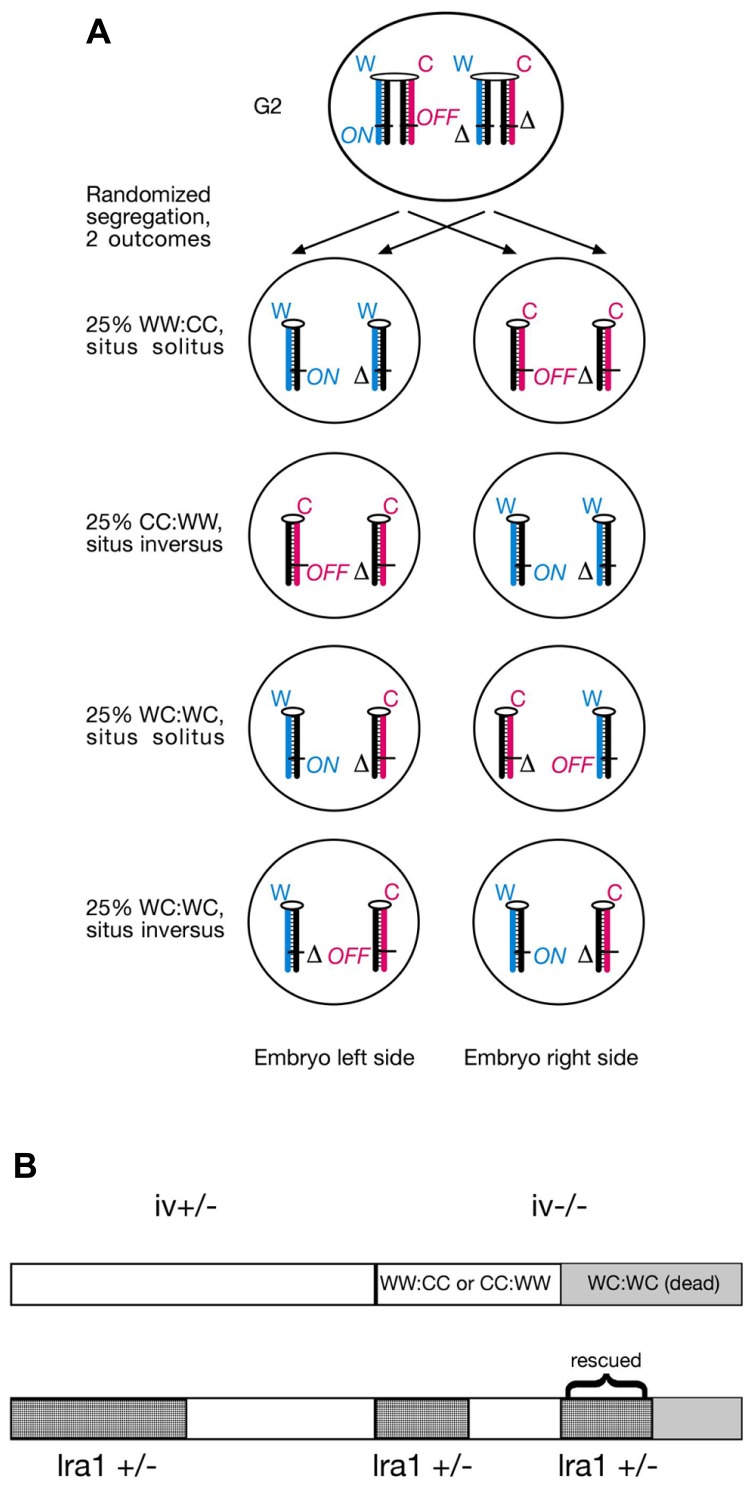
Ever since cloning the classic iv (inversedviscerum) mutation identified the "left-right dynein" (lrd) gene in mice, most research on body laterality determination has focused on its function in motile cilia at the node embryonic organizer. This model is attractive, as it links chirality of cilia architecture to asymmetry development. However, lrd is also expressed in blastocysts and embryonic stem cells, where it was shown to bias the segregation of recombined sister chromatids away from each other in mitosis. These data suggested that lrd is part of a cellular mechanism that recognizes and selectively segregates sister chromatids based on their replication history: old "Watson" versus old "Crick" strands. We previously proposed that the mouse left-right axis is established via an asymmetric cell division prior to/or during gastrulation. In this model, left-right dynein selectively segregates epigenetically differentiated sister chromatids harboring a hypothetical "left-right axis development 1" ("lra1") gene during the left-right axis establishing cell division. Here, asymmetry development would be ultimately governed by the chirality of the cytoskeleton and the DNA molecule. Our model predicts that randomization of chromatid segregation in lrd mutants should produce embryos with 25% situs solitus, 25% situs inversus, and 50% embryonic death due to heterotaxia and isomerism. Here we confirmed this prediction by using two distinct lrd mutant alleles. Other than lrd, thus far Nodal gene is the most upstream function implicated in visceral organs laterality determination. We next tested whether the Nodal gene constitutes the lra1 gene hypothesized in the model by testing mutant's effect on 50% embryonic lethality observed in lrd mutants. Since Nodal mutation did not suppress lethality, we conclude that Nodal is not equivalent to the lra1 gene. In summary, we describe the origin of 50% lethality in lrd mutant mice not yet explained by any other laterality-generating hypothesis.
自从克隆经典的 iv(反向内脏)突变体鉴定出小鼠的“左右动力蛋白”(lrd)基因以来,大多数关于身体左右侧确定的研究都集中在它在节点胚胎组织者中的运动纤毛功能上。这个模型很有吸引力,因为它将纤毛结构的手性与不对称发育联系起来。然而,lrd 也在胚泡和胚胎干细胞中表达,在那里它被证明在有丝分裂中使重组姐妹染色单体偏向彼此分离。这些数据表明,lrd 是一种细胞机制的一部分,该机制根据其复制历史识别并选择性地分离姐妹染色单体:旧的“沃森”与旧的“克里克”链。我们之前提出,小鼠的左右轴是在原肠胚形成之前/期间通过不对称细胞分裂建立的。在这个模型中,左右动力蛋白在建立左右轴的细胞分裂中选择性地分离具有假设的“左右轴发育 1”(“lra1”)基因的表观遗传分化的姐妹染色单体。在这里,不对称发育最终将由细胞骨架和 DNA 分子的手性来控制。我们的模型预测,lrd 突变体中染色单体分离的随机化应该产生 25% situs solitus、25% situs inversus 和 50%由于异位和同型性而导致的胚胎死亡。在这里,我们使用两个不同的 lrd 突变等位基因证实了这一预测。除了 lrd 之外,到目前为止,Nodal 基因是最上游的功能,涉及内脏器官左右侧确定。我们接下来测试了 Nodal 基因是否构成模型中假设的 lra1 基因,方法是测试突变对 lrd 突变体中观察到的 50%胚胎致死的影响。由于 Nodal 突变不能抑制致死性,我们得出结论,Nodal 与 lra1 基因不等同。总之,我们描述了 lrd 突变小鼠中 50%致死率的起源,这一现象尚未被任何其他产生左右侧的假说解释。