Howard Hughes Medical Institute, Harvard University, Cambridge, MA 02138, USA.
Cell. 2013 Jan 31;152(3):442-52. doi: 10.1016/j.cell.2012.12.040.
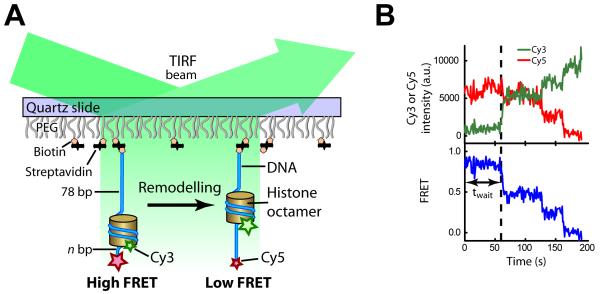
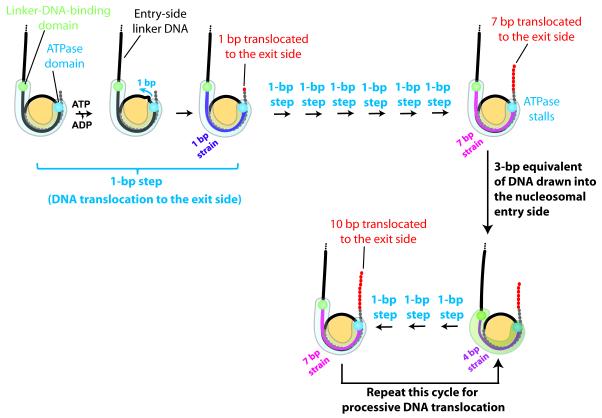
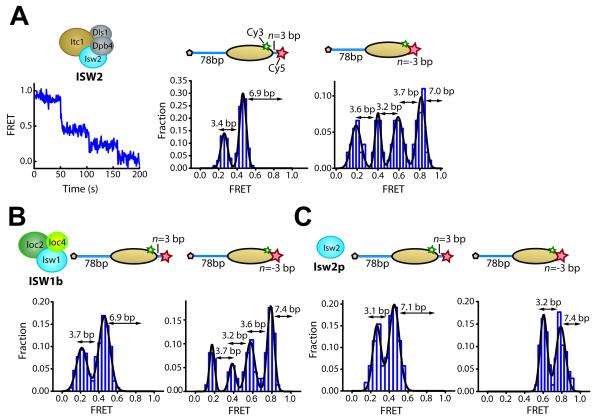
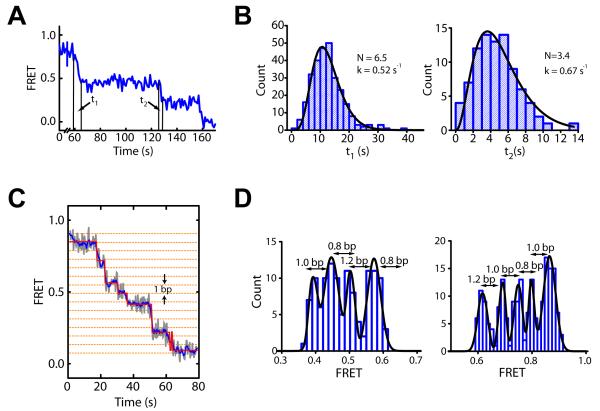
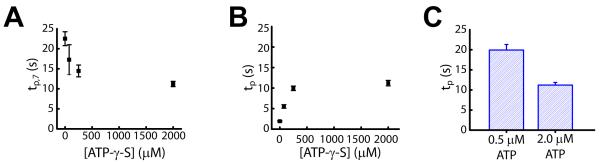
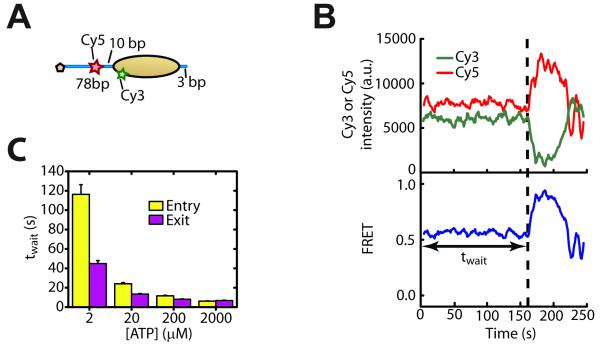
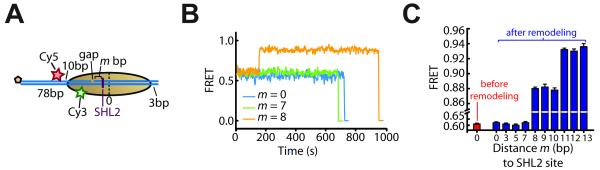
ISWI-family enzymes remodel chromatin by sliding nucleosomes along DNA, but the nucleosome translocation mechanism remains unclear. Here we use single-molecule FRET to probe nucleosome translocation by ISWI-family remodelers. Distinct ISWI-family members translocate nucleosomes with a similar stepping pattern maintained by the catalytic subunit of the enzyme. Nucleosome remodeling begins with a 7 bp step of DNA translocation followed by 3 bp subsequent steps toward the exit side of nucleosomes. These multi-bp, compound steps are comprised of 1 bp substeps. DNA movement on the entry side of the nucleosome occurs only after 7 bp of exit-side translocation, and each entry-side step draws in a 3 bp equivalent of DNA that allows three additional base pairs to be moved to the exit side. Our results suggest a remodeling mechanism with well-defined coordination at different nucleosomal sites featuring DNA translocation toward the exit side in 1 bp steps preceding multi-bp steps of DNA movement on the entry side.
ISWI 家族酶通过沿 DNA 滑动核小体来重塑染色质,但核小体易位机制仍不清楚。在这里,我们使用单分子 FRET 来探测 ISWI 家族重塑酶的核小体易位。不同的 ISWI 家族成员通过酶的催化亚基以相似的步进模式转运核小体。核小体重塑首先是 DNA 易位的 7 bp 步,然后是朝向核小体出口侧的 3 bp 后续步骤。这些多 bp 的复合步骤由 1 bp 的亚步骤组成。核小体进入侧的 DNA 运动仅在出口侧的 7 bp 易位后发生,并且每个进入侧步骤都引入相当于 3 bp 的 DNA,从而允许另外三个碱基对移动到出口侧。我们的结果表明,在不同核小体位点具有明确协调的重塑机制,其特征在于朝向出口侧的 DNA 易位以 1 bp 步为先导,随后是进入侧的 DNA 运动的多 bp 步。