Centre of Microbial and Plant Genetics, Department of Microbial and Molecular Systems, University of Leuven, Heverlee-Leuven, Belgium.
PLoS One. 2013 May 17;8(5):e62946. doi: 10.1371/journal.pone.0062946. Print 2013.
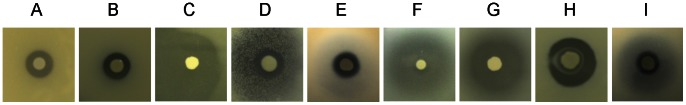
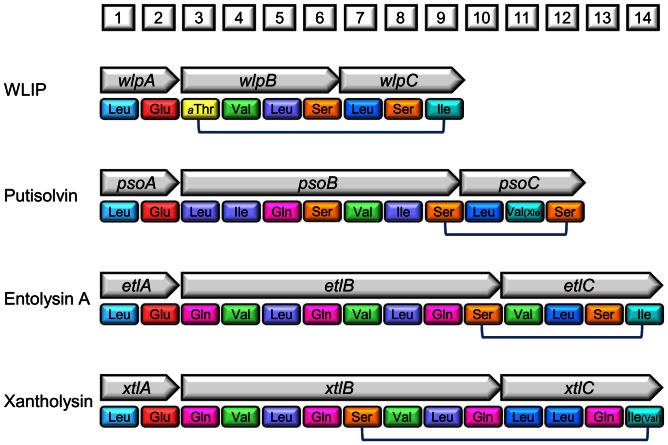
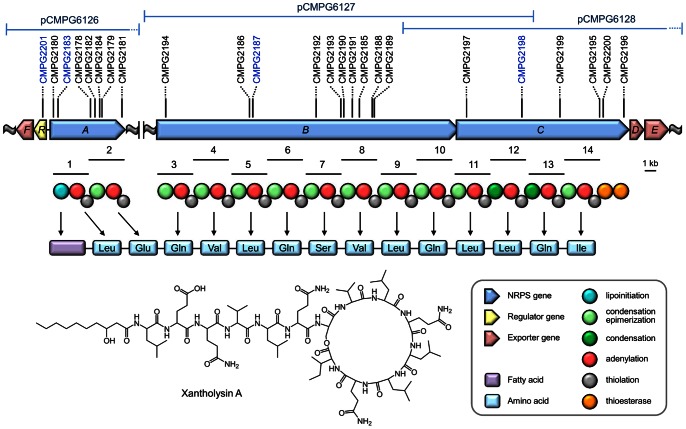
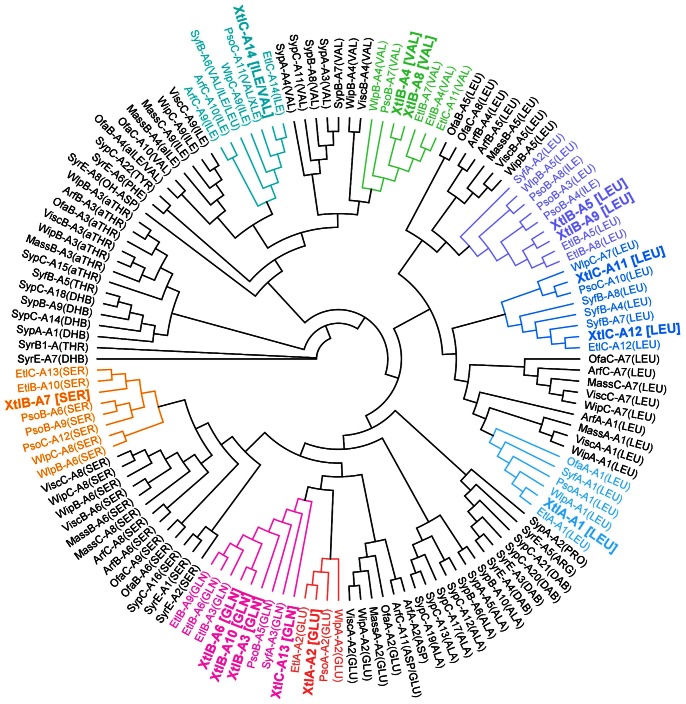
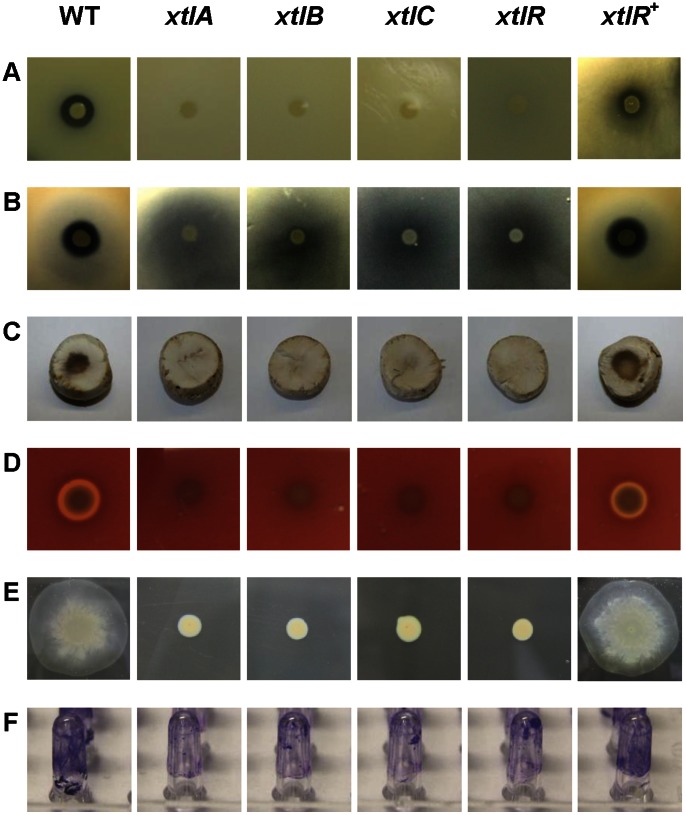
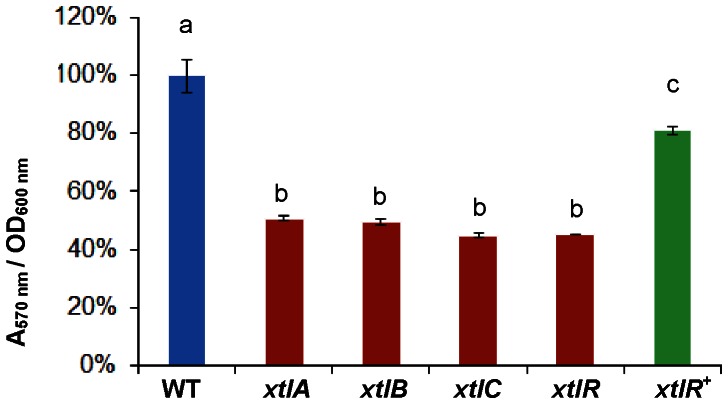
The rhizosphere isolate Pseudomonas putida BW11M1 produces a mixture of cyclic lipopeptide congeners, designated xantholysins. Properties of the major compound xantholysin A, shared with several other Pseudomonas lipopeptides, include antifungal activity and toxicity to Gram-positive bacteria, a supportive role in biofilm formation, and facilitation of surface colonization through swarming. Atypical is the lipopeptide's capacity to inhibit some Gram-negative bacteria, including several xanthomonads. The lipotetradecadepsipeptides are assembled by XtlA, XtlB and XtlC, three co-linearly operating non-ribosomal peptide synthetases (NRPSs) displaying similarity in modular architecture with the entolysin-producing enzymes of the entomopathogenic Pseudomonas entomophila L48. A shifted serine-incorporating unit in the eight-module enzyme XtlB elongating the central peptide moiety not only generates an amino acid sequence differing at several equivalent positions from entolysin, but also directs xantholysin's macrocyclization into an octacyclic structure, distinct from the pentacyclic closure in entolysin. Relaxed fatty acid specificity during lipoinitiation by XtlA (acylation with 3-hydroxydodec-5-enoate instead of 3-hydroxydecanoate) and for incorporation of the ultimate amino acid by XtlC (valine instead of isoleucine) account for the production of the minor structural variants xantholysin C and B, respectively. Remarkably, the genetic backbones of the xantholysin and entolysin NRPS systems also bear pronounced phylogenetic similarity to those of the P. putida strains PCL1445 and RW10S2, albeit generating the seemingly structurally unrelated cyclic lipopeptides putisolvin (undecapeptide containing a cyclotetrapeptide) and WLIP (nonapeptide containing a cycloheptapeptide), respectively. This similarity includes the linked genes encoding the cognate LuxR-family regulator and tripartite export system components in addition to individual modules of the NRPS enzymes, and probably reflects a common evolutionary origin. Phylogenetic scrutiny of the modules used for selective amino acid activation by these synthetases indicates that bacteria such as pseudomonads recruit and reshuffle individual biosynthetic units and blocks thereof to engineer reorganized or novel NRPS assembly lines for diversified synthesis of lipopeptides.
根际分离菌恶臭假单胞菌 BW11M1 产生了一组环脂肽同系物,称为黄杆菌素。主要化合物黄杆菌素 A 的特性与其他几种假单胞菌脂肽共享,包括抗真菌活性和对革兰氏阳性菌的毒性、生物膜形成的支持作用以及通过群集促进表面定植。不同寻常的是,脂肽能够抑制包括几种黄单胞菌在内的一些革兰氏阴性菌。这些脂十四肽是由 XtlA、XtlB 和 XtlC 组装而成的,这三种非核糖体肽合成酶(NRPS)在模块结构上与昆虫病原假单胞菌 L48 的肠毒素产生酶具有相似性。在延长中央肽部分的八模块酶 XtlB 中,掺入丝氨酸的单元不仅在几个等效位置产生了与肠毒素不同的氨基酸序列,而且还指导黄杆菌素的大环化形成八元环结构,与肠毒素中的五元环封闭不同。XtlA 在脂肪酰基起始时对脂肪酸特异性的放松(用 3-羟基十二烷-5-烯酸而不是 3-羟基癸酸酰化)以及 XtlC 对最终氨基酸的掺入(缬氨酸而不是异亮氨酸)分别解释了结构变体黄杆菌素 C 和 B 的产生。值得注意的是,黄杆菌素和肠毒素 NRPS 系统的遗传骨架与 P. putida 菌株 PCL1445 和 RW10S2 的遗传骨架也具有明显的系统发育相似性,尽管它们分别产生看似结构上不相关的环状脂肽 putisolvin(包含环四肽的十一肽)和 WLIP(包含环庚肽的九肽)。这种相似性包括编码同源 LuxR 家族调节剂和三部分出口系统组件的连锁基因,以及 NRPS 酶的单个模块,可能反映了共同的进化起源。对这些合成酶用于选择性氨基酸激活的模块的系统发育分析表明,假单胞菌等细菌会招募和重新排列单个生物合成单元及其模块,以构建重组或新型 NRPS 组装线,用于多样化的脂肽合成。