Department of Earth Sciences, University College London, London, United Kingdom.
PLoS One. 2013 May 15;8(5):e63115. doi: 10.1371/journal.pone.0063115. Print 2013.
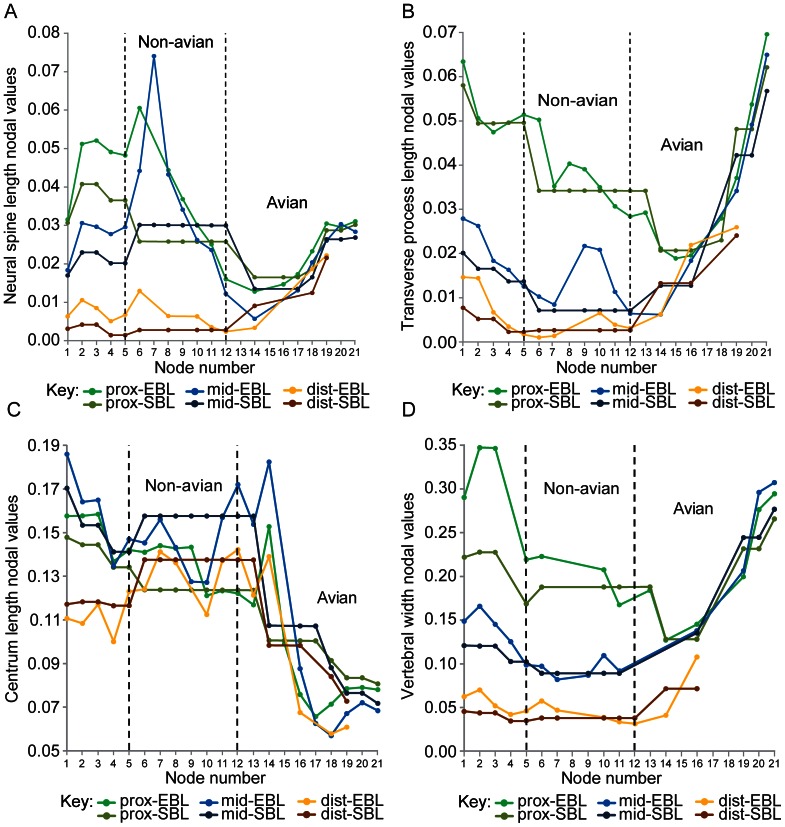
Theropod dinosaurs show striking morphological and functional tail variation; e.g., a long, robust, basal theropod tail used for counterbalance, or a short, modern avian tail used as an aerodynamic surface. We used a quantitative morphological and functional analysis to reconstruct intervertebral joint stiffness in the tail along the theropod lineage to extant birds. This provides new details of the tail's morphological transformation, and for the first time quantitatively evaluates its biomechanical consequences. We observe that both dorsoventral and lateral joint stiffness decreased along the non-avian theropod lineage (between nodes Theropoda and Paraves). Our results show how the tail structure of non-avian theropods was mechanically appropriate for holding itself up against gravity and maintaining passive balance. However, as dorsoventral and lateral joint stiffness decreased, the tail may have become more effective for dynamically maintaining balance. This supports our hypothesis of a reduction of dorsoventral and lateral joint stiffness in shorter tails. Along the avian theropod lineage (Avialae to crown group birds), dorsoventral and lateral joint stiffness increased overall, which appears to contradict our null expectation. We infer that this departure in joint stiffness is specific to the tail's aerodynamic role and the functional constraints imposed by it. Increased dorsoventral and lateral joint stiffness may have facilitated a gradually improved capacity to lift, depress, and swing the tail. The associated morphological changes should have resulted in a tail capable of producing larger muscular forces to utilise larger lift forces in flight. Improved joint mobility in neornithine birds potentially permitted an increase in the range of lift force vector orientations, which might have improved flight proficiency and manoeuvrability. The tail morphology of modern birds with tail fanning capabilities originated in early ornithuromorph birds. Hence, these capabilities should have been present in the early Cretaceous, with incipient tail-fanning capacity in the earliest pygostylian birds.
兽脚亚目恐龙的尾巴在形态和功能上表现出显著的变化,例如,长而强壮的基础兽脚亚目尾巴用于平衡,或短而现代的鸟类尾巴用作空气动力学表面。我们使用定量形态和功能分析来重建沿兽脚亚目谱系到现存鸟类的尾巴椎间关节刚度。这提供了尾巴形态变化的新细节,并首次定量评估了其生物力学后果。我们观察到,在非鸟兽脚亚目恐龙谱系中(在节点兽脚亚目和副爬行动物之间),背腹侧和侧向关节刚度都降低了。我们的结果表明,非鸟兽脚亚目恐龙的尾巴结构在机械上适合支撑自身重力并保持被动平衡。然而,随着背腹侧和侧向关节刚度的降低,尾巴可能更有效地用于动态维持平衡。这支持了我们关于缩短尾巴时减少背腹侧和侧向关节刚度的假设。沿着鸟类兽脚亚目谱系(从鸟兽到冠群鸟类),背腹侧和侧向关节刚度总体上增加了,这似乎与我们的零假设相矛盾。我们推断,这种关节刚度的偏离是尾巴的空气动力学作用和由此产生的功能约束所特有的。增加的背腹侧和侧向关节刚度可能有助于逐渐提高提升、降低和摆动尾巴的能力。相关的形态变化应该导致尾巴能够产生更大的肌肉力量,以利用飞行中的更大升力。新鸟类中关节灵活性的提高可能允许增加升力向量方向的范围,这可能提高了飞行的熟练程度和机动性。具有扇形尾巴能力的现代鸟类的尾巴起源于早期的鸟类。因此,这些能力应该在早白垩世就存在,最早的尾综骨鸟类就具有初步的扇形尾巴能力。