Department of Chemistry and Biochemistry, Institute for Cellular and Molecular Biology, University of Texas at Austin, Austin, TX 78712, USA.
J Mol Biol. 2013 Aug 9;425(15):2670-86. doi: 10.1016/j.jmb.2013.05.008. Epub 2013 May 20.
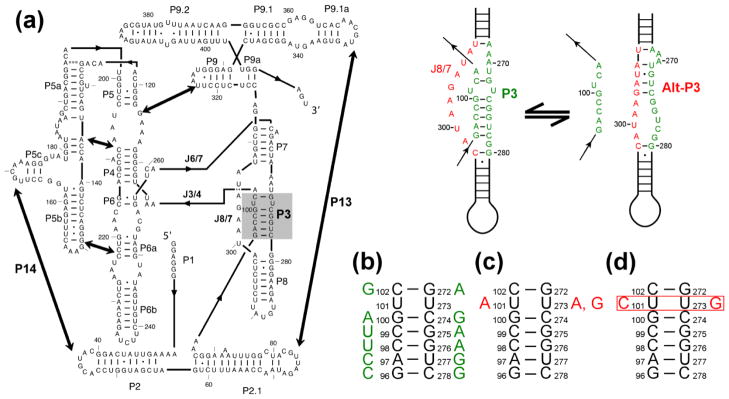
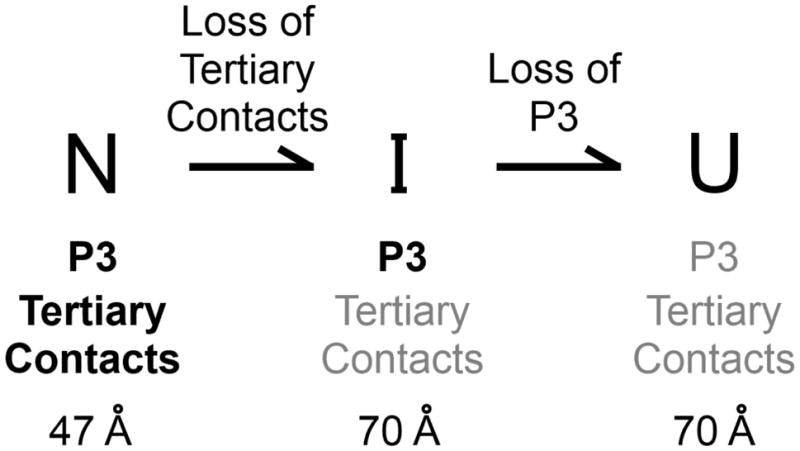
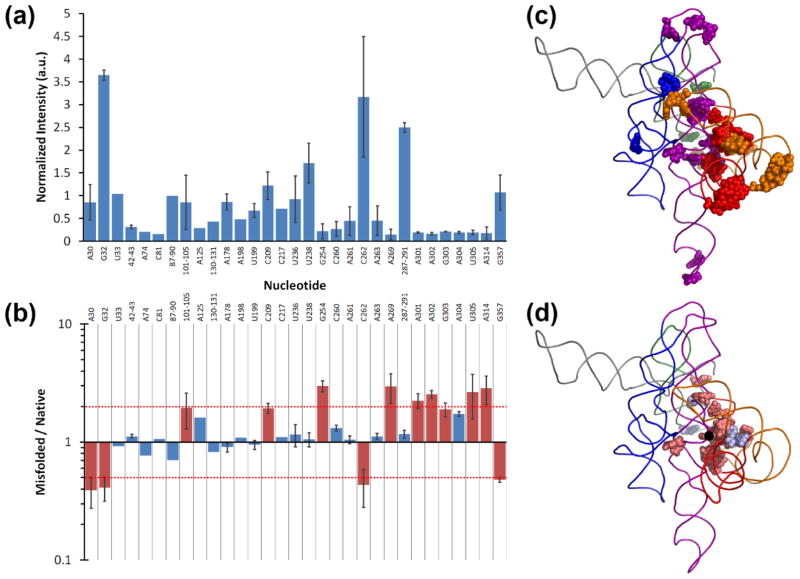
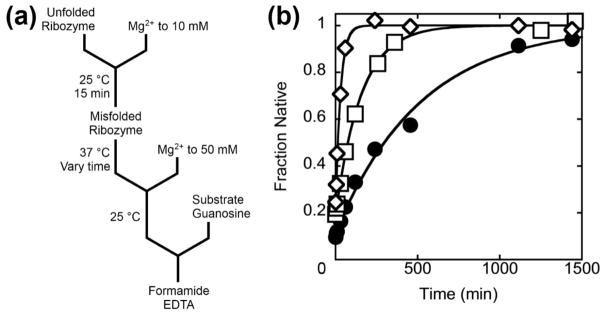
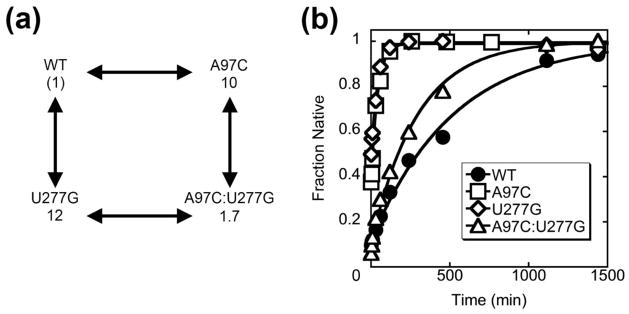
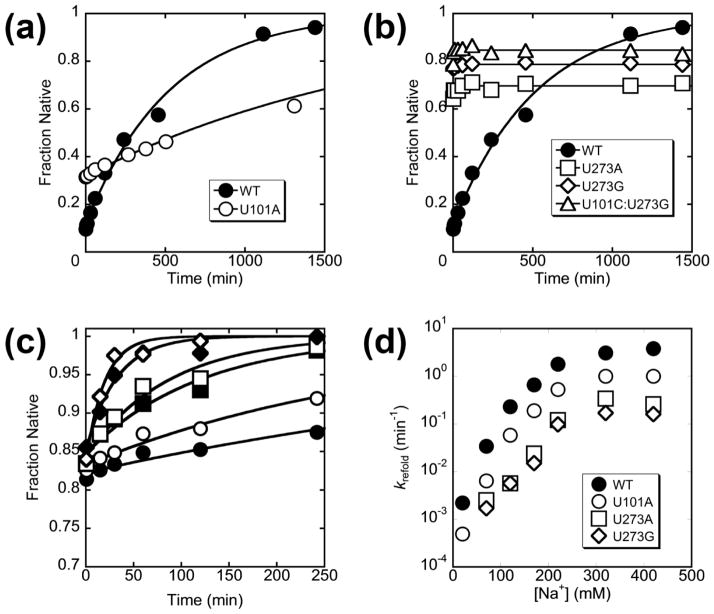
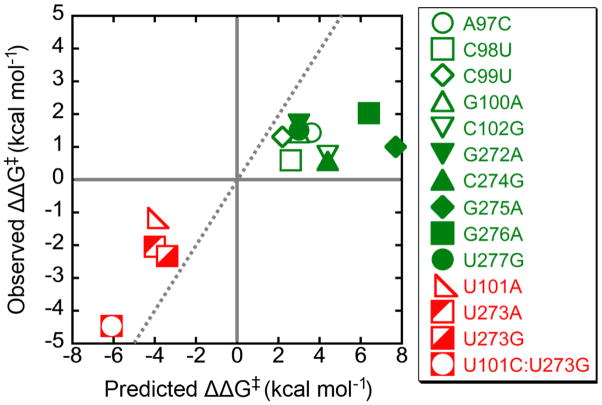
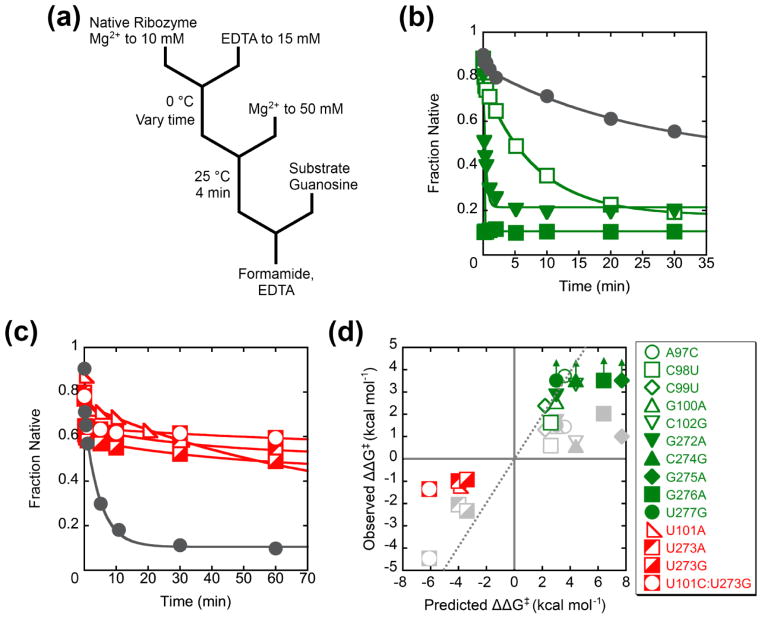
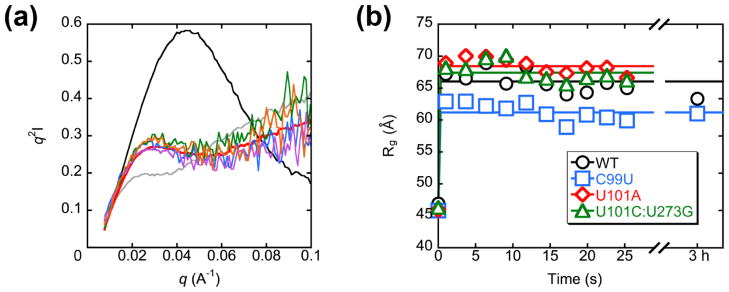
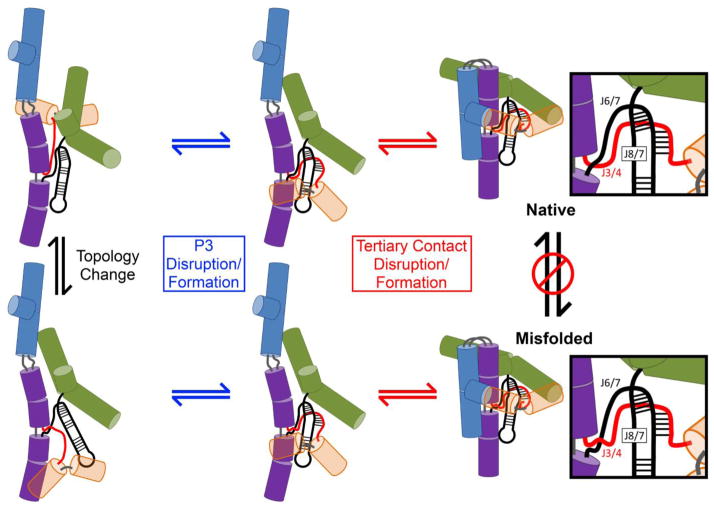
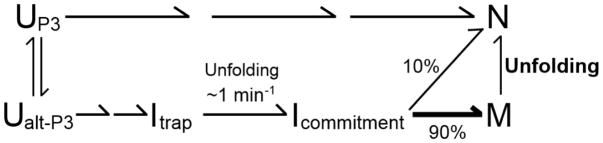
RNAs are prone to misfolding, but how misfolded structures are formed and resolved remains incompletely understood. The Tetrahymena group I intron ribozyme folds in vitro to a long-lived misfolded conformation (M) that includes extensive native structure but is proposed to differ in topology from the native state (N). A leading model predicts that exchange of the topologies requires unwinding of the long-range, core helix P3, despite the presence of P3 in both conformations. To test this model, we constructed 16 mutations to strengthen or weaken P3. Catalytic activity and in-line probing showed that nearly all of the mutants form the M state before folding to N. The P3-weakening mutations accelerated refolding from M (3- to 30-fold) and the P3-strengthening mutations slowed refolding (6- to 1400-fold), suggesting that P3 indeed unwinds transiently. Upon depletion of Mg(2+), the mutations had analogous effects on unfolding from N to intermediates that subsequently fold to M. The magnitudes for the P3-weakening mutations were larger than in refolding from M, and small-angle X-ray scattering showed that the ribozyme expands rapidly to intermediates from which P3 is disrupted subsequently. These results are consistent with previous results indicating unfolding of native peripheral structure during refolding from M, which probably permits rearrangement of the core. Together, our results demonstrate that exchange of the native and misfolded conformations requires loss of a core helix in addition to peripheral structure. Further, the results strongly suggest that misfolding arises from a topological error within the ribozyme core, and a specific topology is proposed.
RNAs 容易发生错误折叠,但错误折叠结构是如何形成和解析的仍不完全清楚。四膜虫 I 类内含子核酶在体外折叠成一种长寿命的错误折叠构象(M),它包含广泛的天然结构,但被认为在拓扑结构上与天然状态(N)不同。一个主要的模型预测,拓扑结构的交换需要解开远距离的核心螺旋 P3,尽管 P3 存在于两种构象中。为了验证这个模型,我们构建了 16 个突变来增强或削弱 P3。催化活性和在线探测表明,几乎所有的突变体在折叠到 N 之前都形成了 M 状态。P3 弱化突变加速了从 M 到 N 的重折叠(3 到 30 倍),而 P3 强化突变减缓了重折叠(6 到 1400 倍),这表明 P3 确实会短暂解开。当 Mg(2+)耗尽时,这些突变对从 N 到中间体的解折叠具有类似的影响,随后中间体折叠到 M。P3 弱化突变的幅度大于从 M 重折叠,小角度 X 射线散射表明核酶迅速扩展到中间体,随后 P3 被破坏。这些结果与之前的结果一致,表明在从 M 重折叠过程中,天然外围结构的解折叠可能允许核心的重新排列。总的来说,我们的结果表明,除了外围结构外,交换天然和错误折叠构象还需要丢失核心螺旋。此外,结果强烈表明错误折叠是由核酶核心中的拓扑错误引起的,并提出了一种特定的拓扑结构。