Ludwig Institute for Cancer Research and Department of Cellular and Molecular Medicine, University of California at San Diego, La Jolla, California 92093, USA.
Nat Cell Biol. 2013 Sep;15(9):1056-66. doi: 10.1038/ncb2805. Epub 2013 Jul 21.
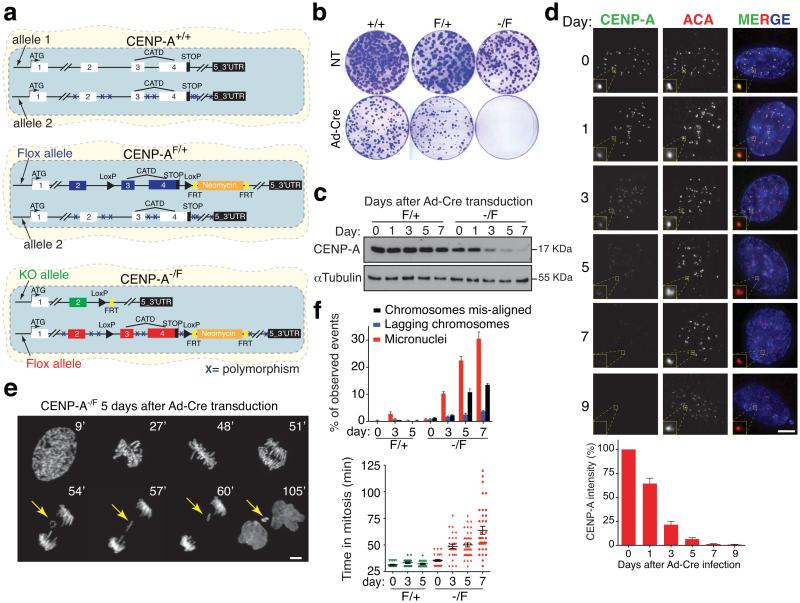
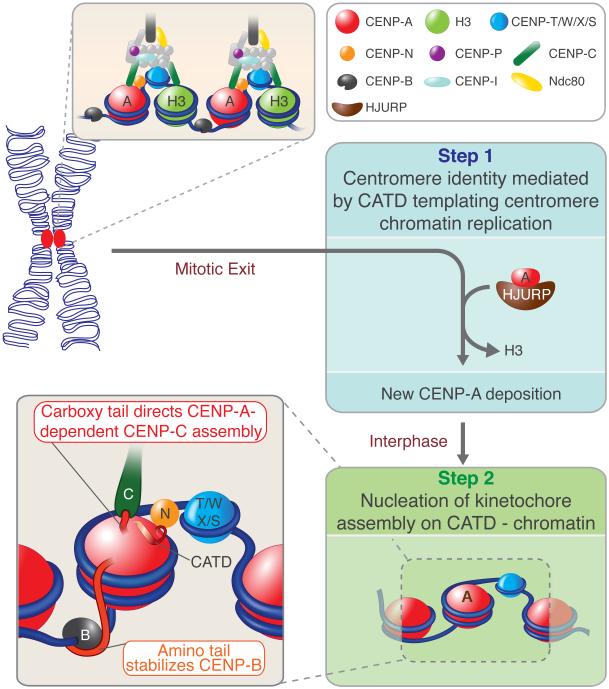
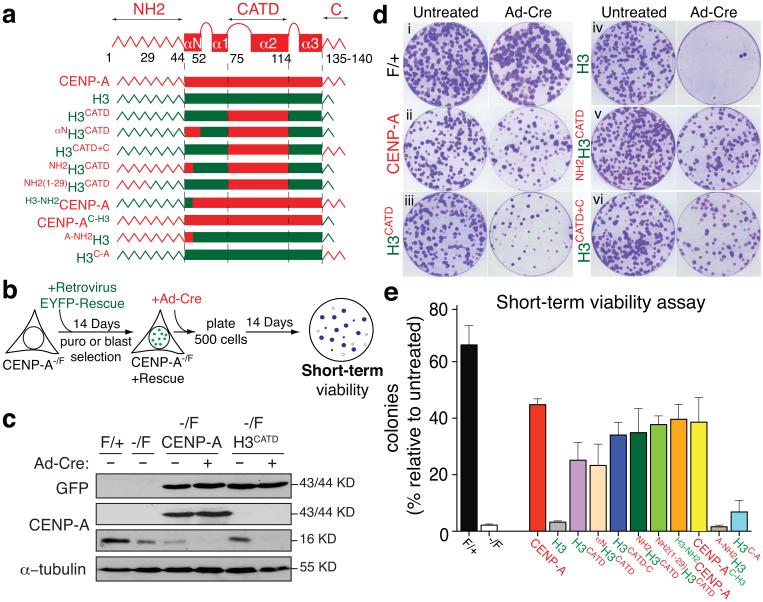
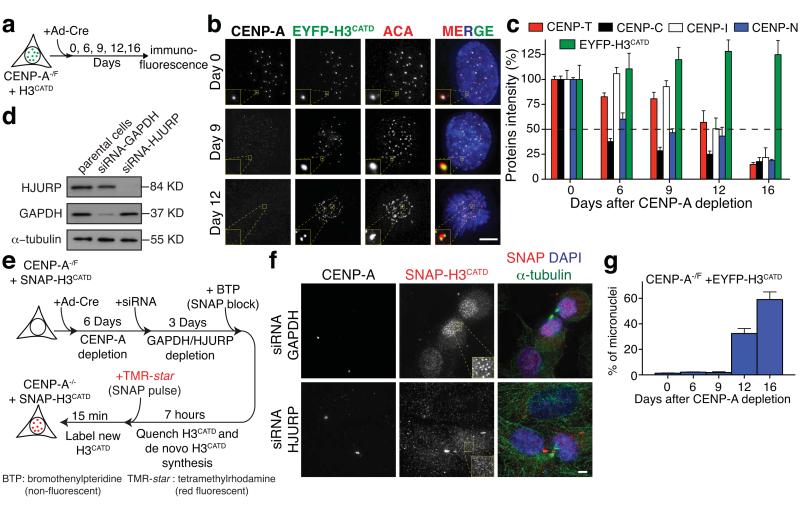
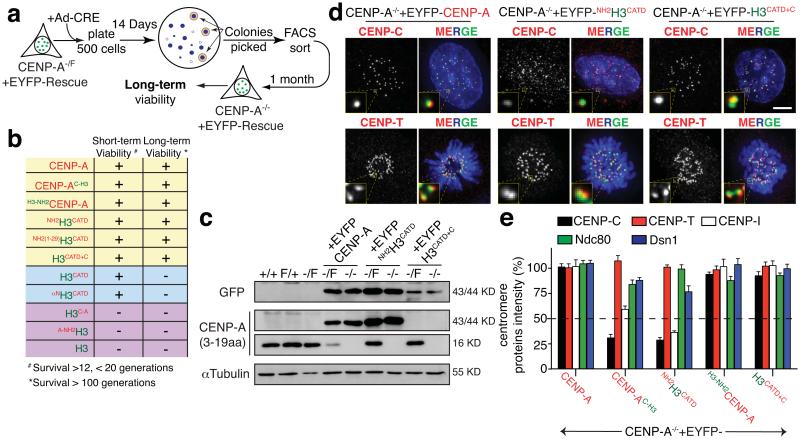
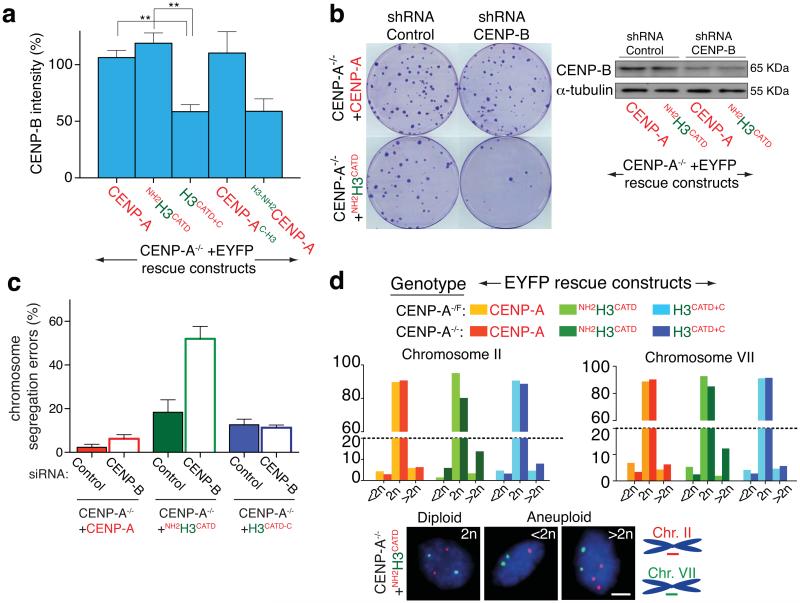
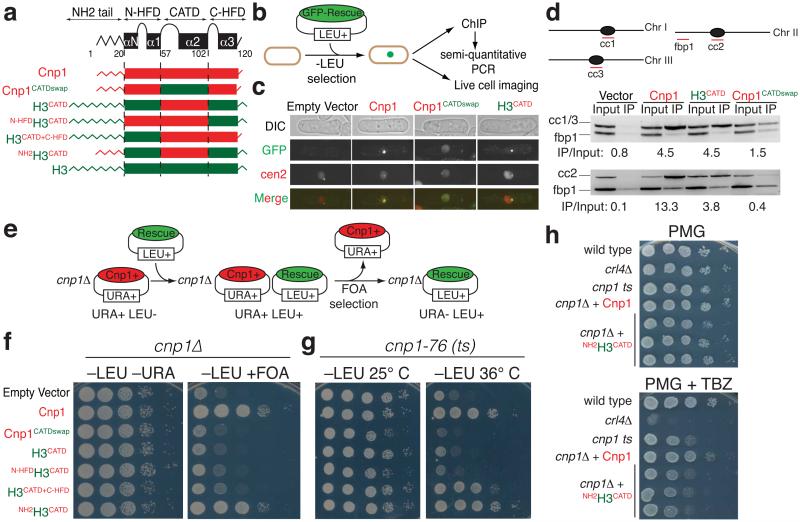
The basic determinant of chromosome inheritance, the centromere, is specified in many eukaryotes by an epigenetic mark. Using gene targeting in human cells and fission yeast, chromatin containing the centromere-specific histone H3 variant CENP-A is demonstrated to be the epigenetic mark that acts through a two-step mechanism to identify, maintain and propagate centromere function indefinitely. Initially, centromere position is replicated and maintained by chromatin assembled with the centromere-targeting domain (CATD) of CENP-A substituted into H3. Subsequently, nucleation of kinetochore assembly onto CATD-containing chromatin is shown to require either the amino- or carboxy-terminal tail of CENP-A for recruitment of inner kinetochore proteins, including stabilizing CENP-B binding to human centromeres or direct recruitment of CENP-C, respectively.
染色体遗传的基本决定因素是着丝粒,在许多真核生物中,着丝粒是由一种表观遗传标记指定的。通过人类细胞和裂殖酵母中的基因靶向,含有着丝粒特异性组蛋白 H3 变体 CENP-A 的染色质被证明是表观遗传标记,通过两步机制来识别、维持和无限期地传播着丝粒功能。最初,通过用 CENP-A 的着丝粒靶向结构域 (CATD) 替代 H3 组装的染色质来复制和维持着丝粒位置。随后,显示出动粒组装的起始需要 CENP-A 的氨基或羧基末端尾巴来招募内动粒蛋白,包括稳定 CENP-B 与人类着丝粒的结合或直接招募 CENP-C。