DaSilva Lance F, Pillon Samantha, Genereaux Julie, Davey Megan J, Gloor Gregory B, Karagiannis Jim, Brandl Christopher J
Department of Biochemistry, Schulich School of Medicine & Dentistry, The University of Western Ontario, London, Canada N6A5C1.
G3 (Bethesda). 2013 Oct 3;3(10):1661-74. doi: 10.1534/g3.113.006841.
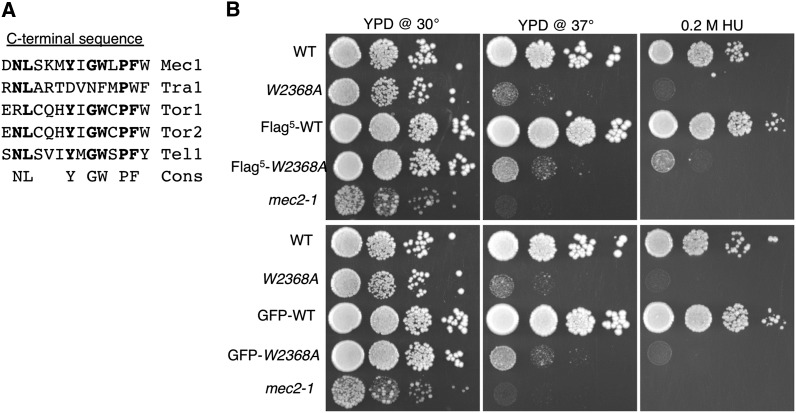
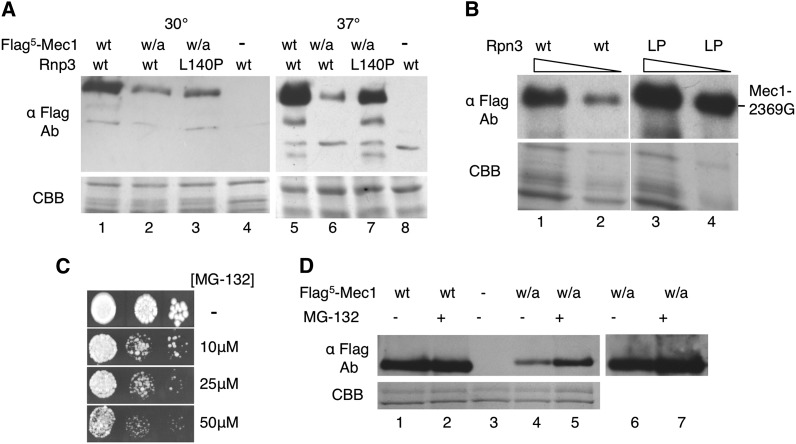
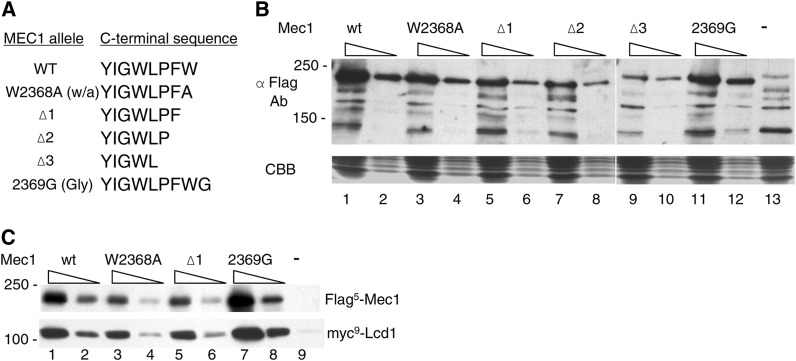
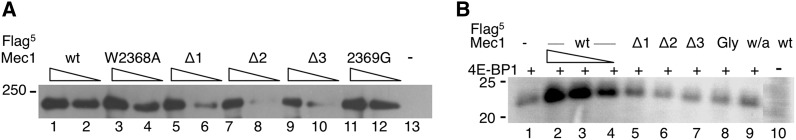
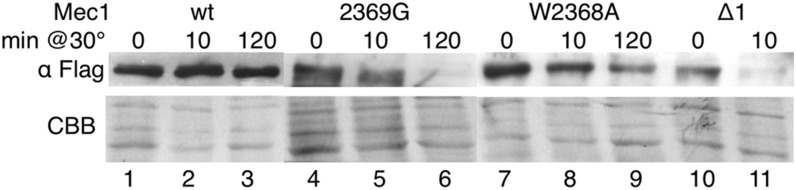
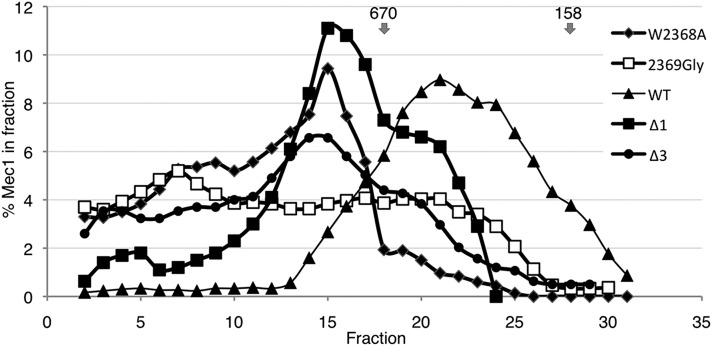
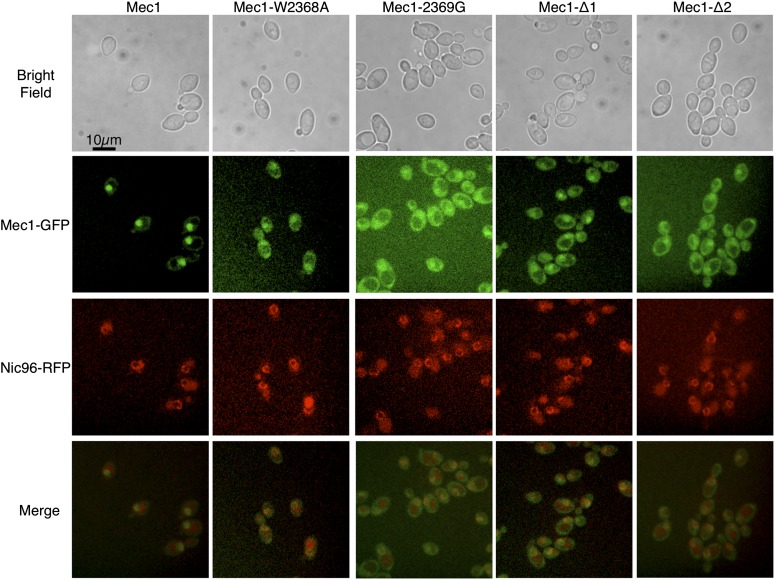
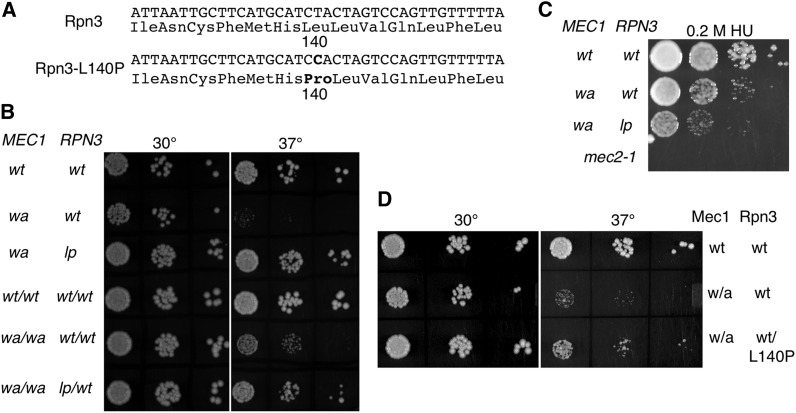
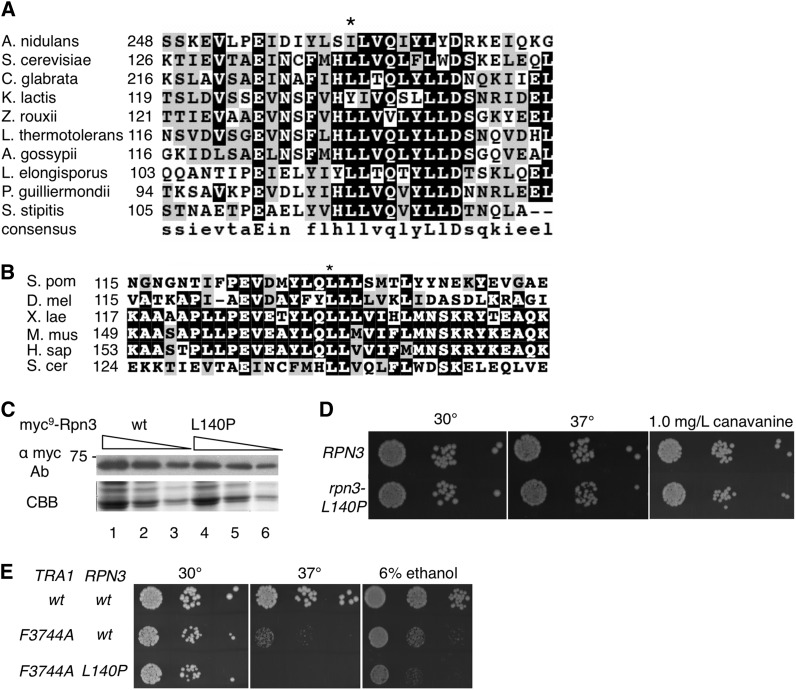
Mec1, a member of the phosphoinositide three-kinase-related kinase (PIKK) family of proteins, is involved in the response to replicative stress and DNA damage and in telomere maintenance. An essential 30 to 35 residue, the FATC domain is found at the C-terminus of all PIKK family members. To investigate the roles of the C-terminal residues of Mec1, we characterized alleles of Saccharomyces cerevisiae mec1 that alter the FATC domain. A change of the terminal tryptophan to alanine resulted in temperature-sensitive growth, sensitivity to hydroxyurea, and diminished kinase activity in vitro. Addition of a terminal glycine or deletion of one, two, or three residues resulted in loss of cell viability and kinase function. Each of these Mec1 derivatives was less stable than wild-type Mec1, eluted abnormally from a size exclusion column, and showed reduced nuclear localization. We identified rpn3-L140P, which encodes a component of the 19S proteasomal regulatory particle of the 26S proteasome, as a suppressor of the temperature-sensitive growth caused by mec1-W2368A. The rpn3-L140P allele acted in a partially dominant fashion. It was not able to suppress the inviability of the C-terminal truncations or additions or the hydroxyurea sensitivity of mec1-W2368A. The rpn3-L140P allele restored Mec1-W2368A to nearly wild-type protein levels at 37°, an effect partially mimicked by the proteasome inhibitor MG-132. Our study supports a role for the C-terminus in Mec1 folding and stability, and suggests a role for the proteasome in regulating Mec1 levels.
Mec1是磷酸肌醇三激酶相关激酶(PIKK)蛋白家族的成员,参与对复制应激和DNA损伤的反应以及端粒维持。FATC结构域是一个由30至35个氨基酸组成的必需结构域,存在于所有PIKK家族成员的C末端。为了研究Mec1 C末端残基的作用,我们对酿酒酵母mec1中改变FATC结构域的等位基因进行了表征。将末端色氨酸突变为丙氨酸导致温度敏感型生长、对羟基脲敏感以及体外激酶活性降低。添加末端甘氨酸或缺失一、二或三个残基导致细胞活力丧失和激酶功能丧失。这些Mec1衍生物中的每一个都比野生型Mec1更不稳定,从尺寸排阻柱上异常洗脱,并显示核定位减少。我们鉴定出rpn3-L140P,它编码26S蛋白酶体19S蛋白酶体调节颗粒的一个组分,是由mec1-W2368A引起的温度敏感型生长的抑制因子。rpn3-L140P等位基因以部分显性方式起作用。它不能抑制C末端截短或添加导致的不可存活性或mec1-W2368A对羟基脲的敏感性。rpn3-L140P等位基因在37°时将Mec1-W2368A恢复到接近野生型的蛋白水平,蛋白酶体抑制剂MG-132可部分模拟这种效果。我们的研究支持C末端在Mec1折叠和稳定性中的作用,并表明蛋白酶体在调节Mec1水平方面的作用。