SynthSys-Synthetic and Systems Biology, The University of Edinburgh, Edinburgh, United Kingdom.
PLoS Comput Biol. 2013;9(8):e1003175. doi: 10.1371/journal.pcbi.1003175. Epub 2013 Aug 8.
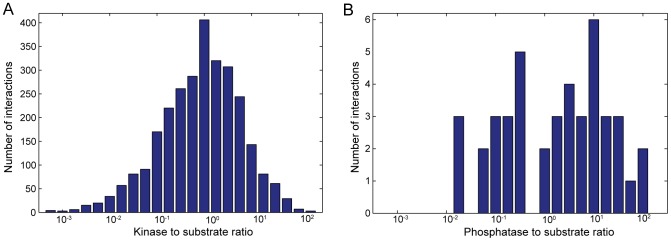
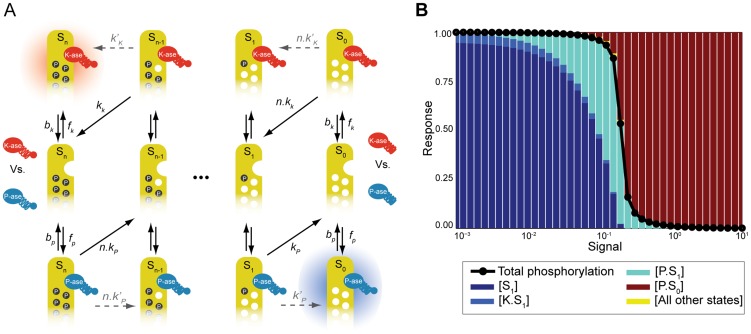
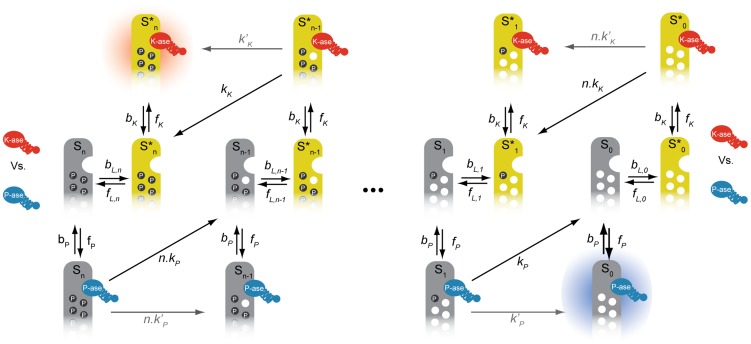
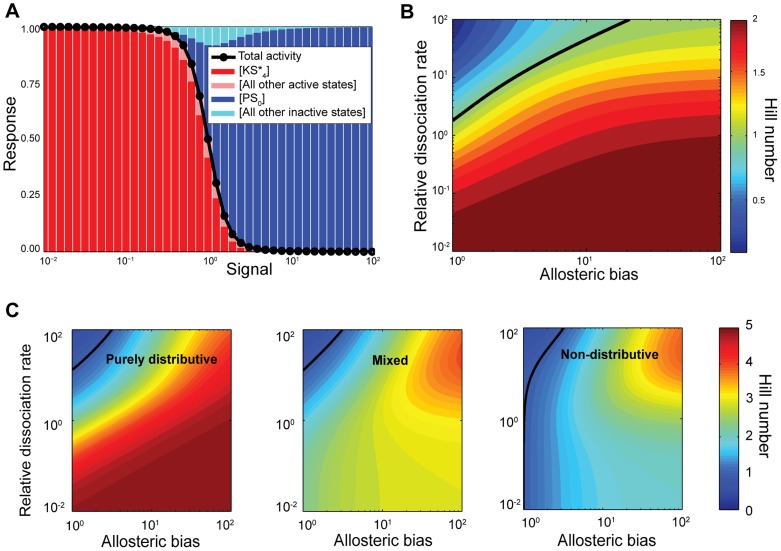
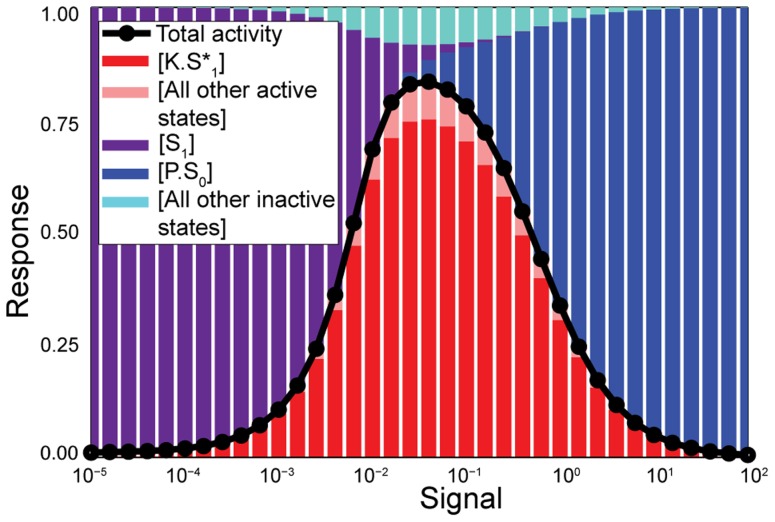
Cellular decision-making is driven by dynamic behaviours, such as the preparations for sunrise enabled by circadian rhythms and the choice of cell fates enabled by positive feedback. Such behaviours are often built upon ultrasensitive responses where a linear change in input generates a sigmoidal change in output. Phosphorylation-dephosphorylation cycles are one means to generate ultrasensitivity. Using bioinformatics, we show that in vivo levels of kinases and phosphatases frequently exceed the levels of their corresponding substrates in budding yeast. This result is in contrast to the conditions often required by zero-order ultrasensitivity, perhaps the most well known means for how such cycles become ultrasensitive. We therefore introduce a mechanism to generate ultrasensitivity when numbers of enzymes are higher than numbers of substrates. Our model combines distributive and non-distributive actions of the enzymes with two-stage binding and concerted allosteric transitions of the substrate. We use analytical and numerical methods to calculate the Hill number of the response. For a substrate with [Formula: see text] phosphosites, we find an upper bound of the Hill number of [Formula: see text], and so even systems with a single phosphosite can be ultrasensitive. Two-stage binding, where an enzyme must first bind to a binding site on the substrate before it can access the substrate's phosphosites, allows the enzymes to sequester the substrate. Such sequestration combined with competition for each phosphosite provides an intuitive explanation for the sigmoidal shifts in levels of phosphorylated substrate. Additionally, we find cases for which the response is not monotonic, but shows instead a peak at intermediate levels of input. Given its generality, we expect the mechanism described by our model to often underlay decision-making circuits in eukaryotic cells.
细胞决策是由动态行为驱动的,例如生物钟驱动的日出准备和正反馈驱动的细胞命运选择。这些行为通常建立在超敏反应的基础上,即输入的线性变化产生输出的阶跃变化。磷酸化-去磷酸化循环是产生超敏反应的一种手段。我们使用生物信息学方法表明,在出芽酵母中,激酶和磷酸酶的体内水平经常超过其相应底物的水平。这一结果与零级超敏反应通常所需的条件形成对比,零级超敏反应可能是此类循环变得超敏的最著名手段。因此,我们引入了一种当酶的数量高于底物的数量时产生超敏反应的机制。我们的模型将酶的分布式和非分布式作用与底物的两步结合和协同变构跃迁结合在一起。我们使用分析和数值方法来计算响应的 Hill 数。对于具有 [Formula: see text] 个磷酸化位点的底物,我们发现响应的 Hill 数的上限为 [Formula: see text],因此即使只有一个磷酸化位点的系统也可以是超敏的。两步结合,其中酶必须首先结合到底物上的一个结合位点,然后才能访问底物的磷酸化位点,使酶能够隔离底物。这种隔离加上对每个磷酸化位点的竞争为磷酸化底物水平的阶跃变化提供了直观的解释。此外,我们还发现了一些情况下,响应不是单调的,而是在中间输入水平出现峰值。鉴于其普遍性,我们预计我们模型所描述的机制通常会成为真核细胞决策电路的基础。