Department of Molecular and Cell Biology, University of California, Berkeley, Berkeley, California, USA.
Nat Struct Mol Biol. 2013 Oct;20(10):1164-72. doi: 10.1038/nsmb.2659. Epub 2013 Sep 8.
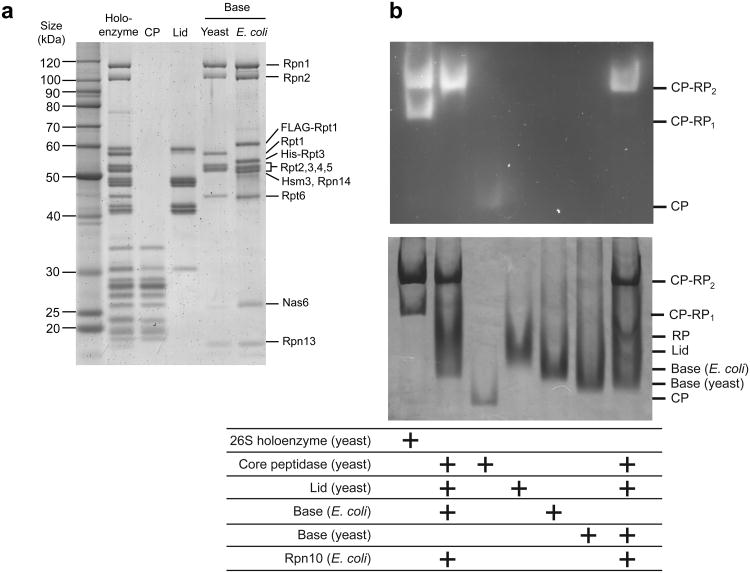
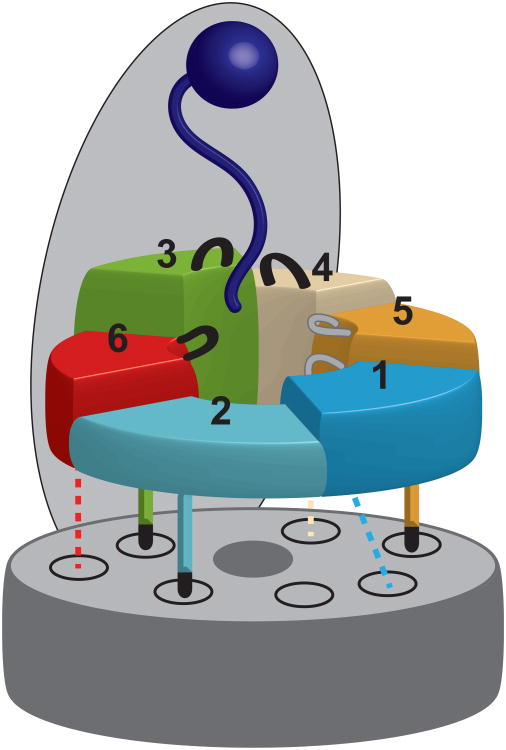
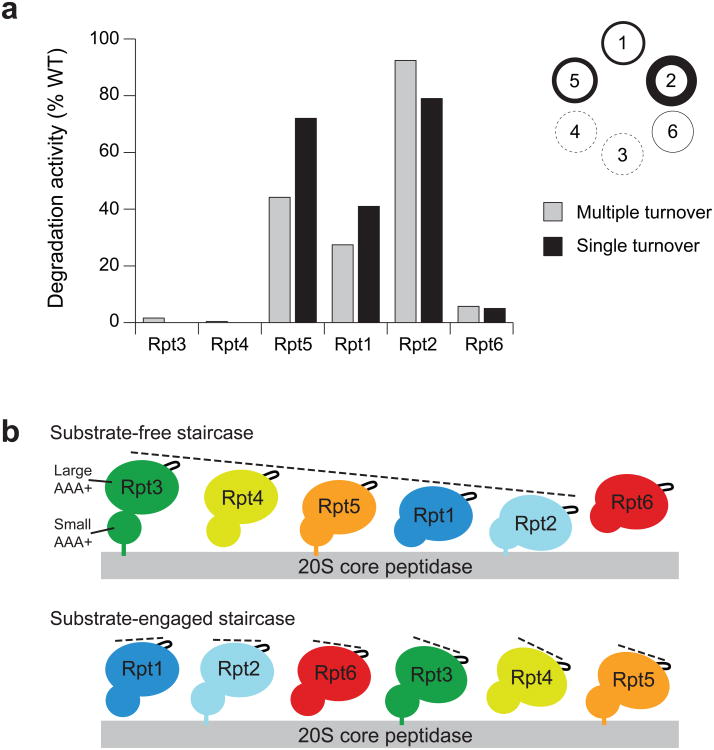
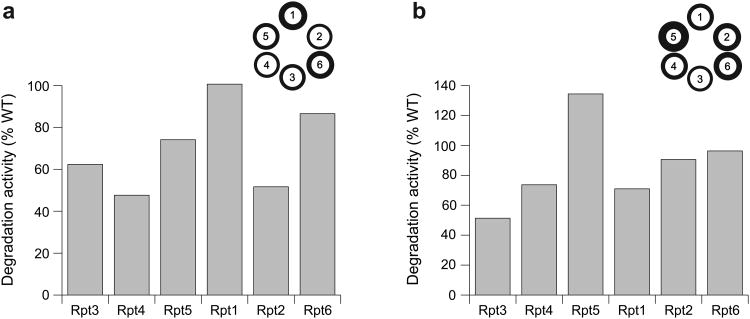
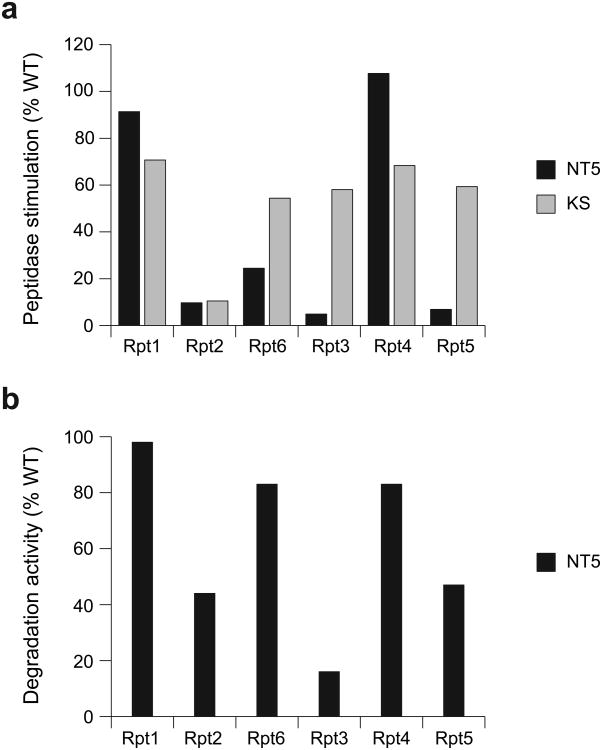
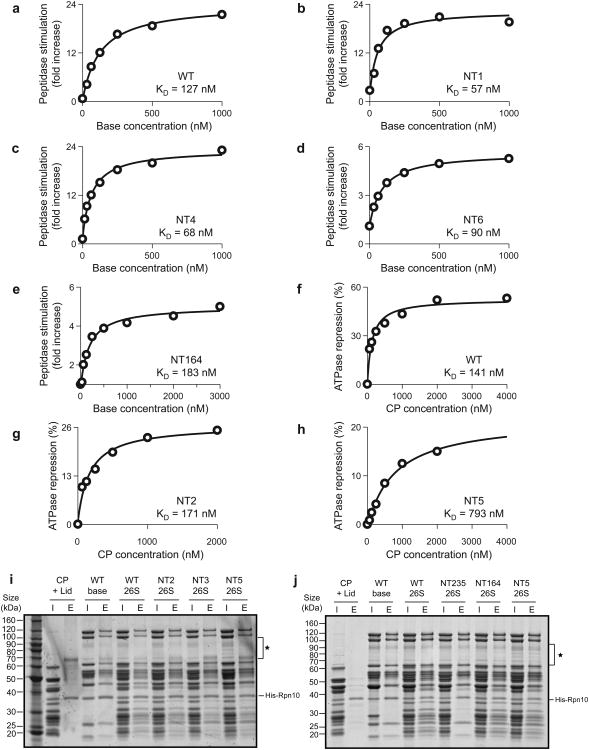
The 26S proteasome is the major eukaryotic ATP-dependent protease, yet the detailed mechanisms used by the proteasomal heterohexameric AAA+ unfoldase to drive substrate degradation remain poorly understood. To perform systematic mutational analyses of individual ATPase subunits, we heterologously expressed the unfoldase subcomplex from Saccharomyces cerevisiae in Escherichia coli and reconstituted the proteasome in vitro. Our studies demonstrate that the six ATPases have distinct roles in degradation, corresponding to their positions in the spiral staircases adopted by the AAA+ domains in the absence or presence of substrate. ATP hydrolysis in subunits at the top of the staircases is critical for substrate engagement and translocation. Whereas the unfoldase relies on this vertical asymmetry for substrate processing, interaction with the peptidase exhibits three-fold symmetry with contributions from alternate subunits. These diverse functional asymmetries highlight how the 26S proteasome deviates from simpler, homomeric AAA+ proteases.
26S 蛋白酶体是主要的真核生物 ATP 依赖性蛋白酶,但蛋白酶体异六聚体 AAA+解旋酶用于驱动底物降解的详细机制仍知之甚少。为了对单个 ATP 酶亚基进行系统的突变分析,我们在大肠杆菌中异源表达了酿酒酵母的解旋酶亚基,并在体外重新组装了蛋白酶体。我们的研究表明,这六个 ATP 酶在降解中具有不同的作用,这与其在 AAA+结构域无底物或有底物时采用的螺旋梯阶中的位置相对应。位于梯级顶部的亚基中的 ATP 水解对于底物结合和转运至关重要。虽然解旋酶依赖这种垂直不对称性来处理底物,但与肽酶的相互作用具有三倍对称性,来自交替的亚基的贡献。这些不同的功能不对称性突出了 26S 蛋白酶体如何偏离更简单的同型 AAA+蛋白酶。