Department of Physiology and Biophysics, University of California, Irvine, Irvine, CA 92697.
J Gen Physiol. 2013 Oct;142(4):405-12. doi: 10.1085/jgp.201311030.
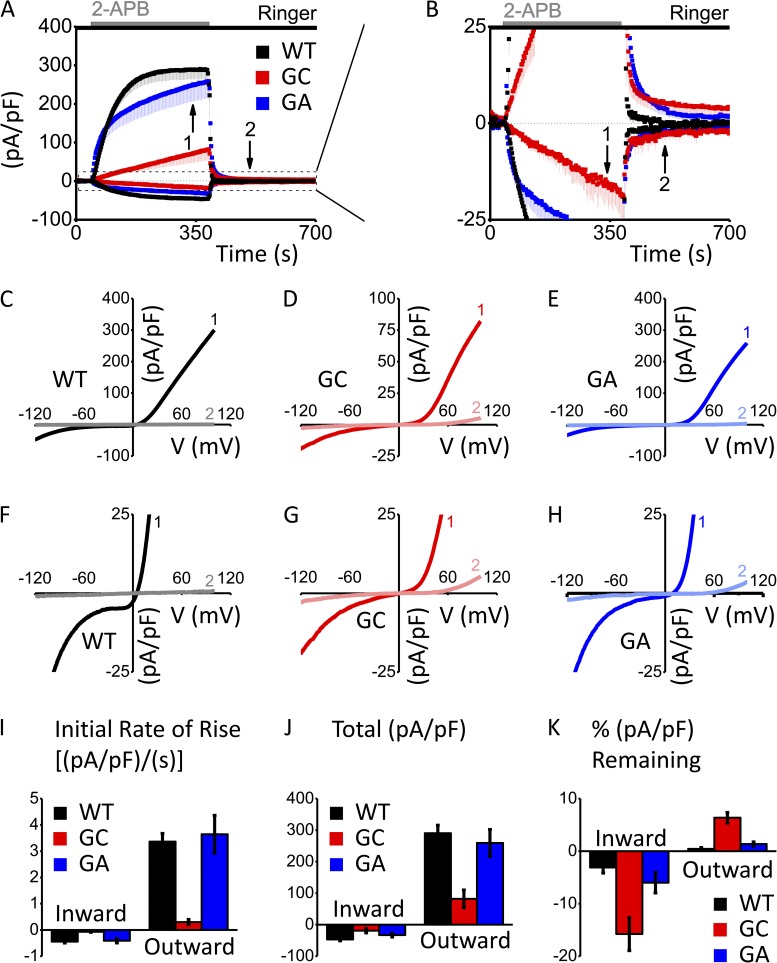
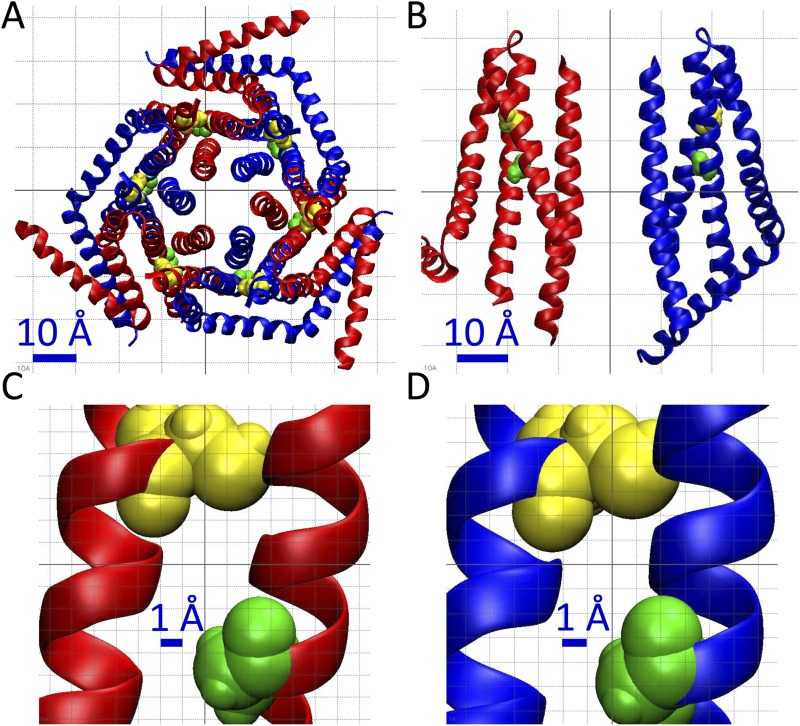
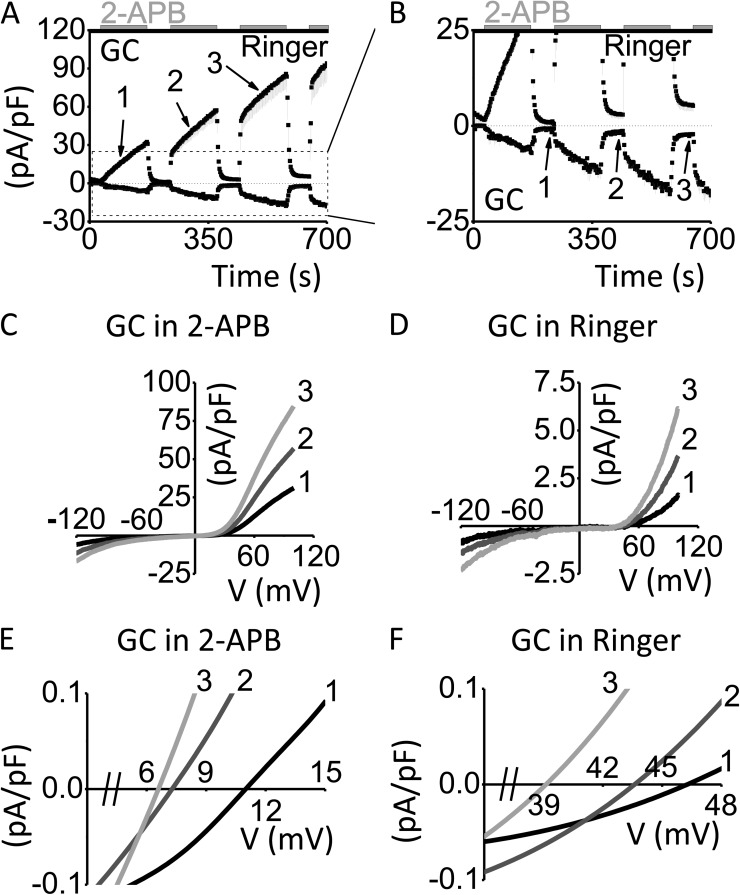
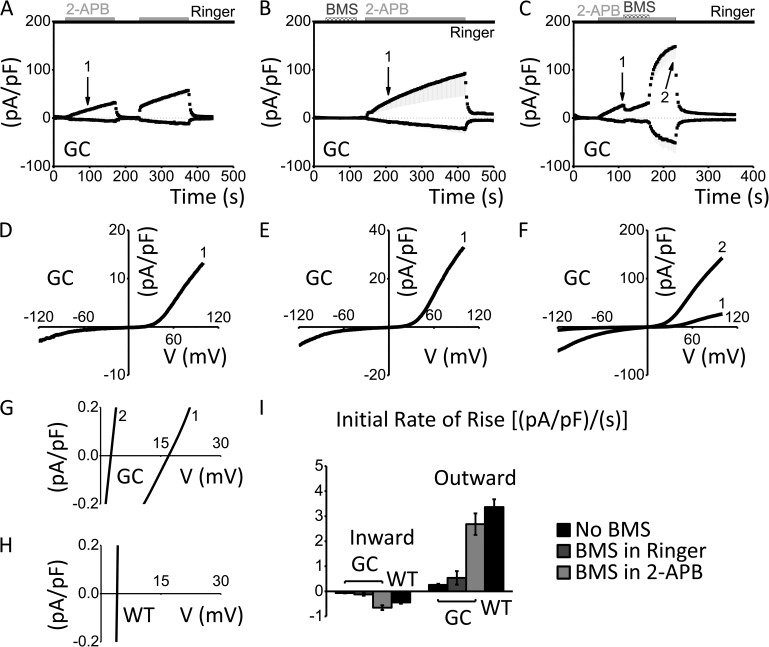
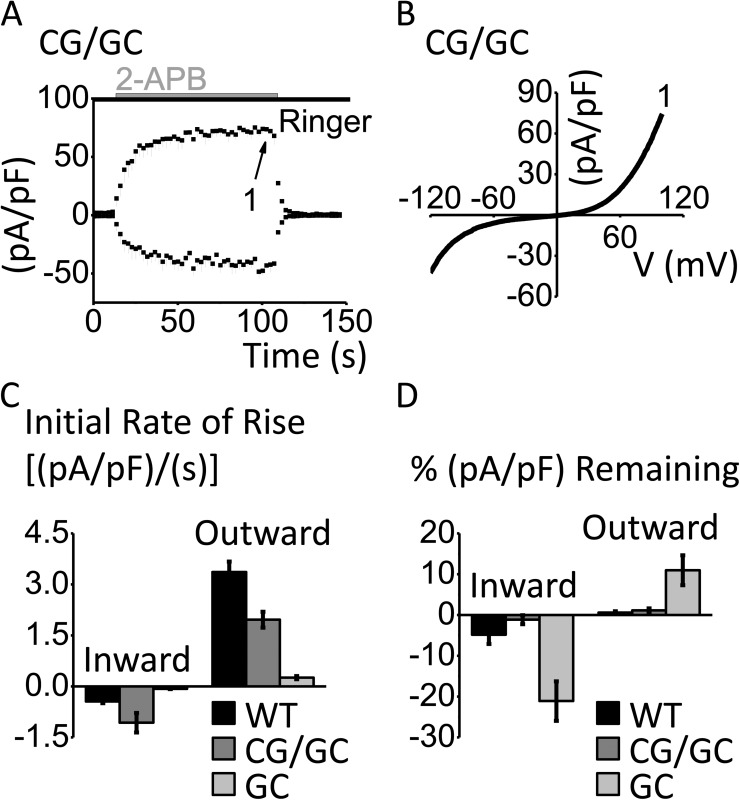
After endoplasmic reticulum (ER) Ca(2+) store depletion, Orai channels in the plasma membrane (PM) are activated directly by ER-resident STIM proteins to form the Ca(2+)-selective Ca(2+) release-activated Ca(2+) (CRAC) channel. However, in the absence of Ca(2+) store depletion and STIM interaction, the mammalian homologue Orai3 can be activated by 2-aminoethyl diphenylborinate (2-APB), resulting in a nonselective cation conductance characterized by biphasic inward and outward rectification. Here, we use site-directed mutagenesis and patch-clamp analysis to better understand the mechanism by which 2-APB activates Orai3. We find that point mutation of glycine 158 in the third transmembrane (TM) segment to cysteine, but not alanine, slows the kinetics of 2-APB activation and prevents complete channel closure upon 2-APB washout. The "slow" phenotype exhibited by Orai3 mutant G158C reveals distinct open states, characterized by variable reversal potentials. The slow phenotype can be reversed by application of the reducing reagent bis(2-mercaptoethylsulfone) (BMS), but in a state-dependent manner, only during 2-APB activation. Moreover, the double mutant C101G/G158C, in which an endogenous TM2 cysteine is changed to glycine, does not exhibit altered kinetics of 2-APB activation. We suggest that a disulfide bridge, formed between the introduced cysteine at TM3 position 158 and the endogenous cysteine at TM2 position 101, hinders transitions between Orai3 open and closed states. Our data provide functional confirmation of the proximity of these two residues and suggest a location within the Orai3 protein that is sensitive to the actions of 2-APB.
内质网 (ER) Ca(2+) 储存耗尽后,质膜 (PM) 中的 Orai 通道被 ER 驻留的 STIM 蛋白直接激活,形成 Ca(2+) 选择性 Ca(2+) 释放激活的 Ca(2+) (CRAC) 通道。然而,在没有 Ca(2+) 储存耗尽和 STIM 相互作用的情况下,哺乳动物同源物 Orai3 可以被 2-氨基乙基二苯基硼酸盐 (2-APB) 激活,导致具有双相内向和外向整流的非选择性阳离子电导。在这里,我们使用定点突变和膜片钳分析来更好地理解 2-APB 激活 Orai3 的机制。我们发现,第三跨膜 (TM) 段中的甘氨酸 158 点突变为半胱氨酸,但不是丙氨酸,会减缓 2-APB 激活的动力学,并防止在 2-APB 冲洗后完全关闭通道。Orai3 突变体 G158C 表现出的“慢”表型揭示了不同的开放状态,其特征是可变的反转电位。“慢”表型可以通过施加还原剂双(2-巯基乙基磺酰) (BMS) 来逆转,但以状态依赖的方式,仅在 2-APB 激活期间。此外,内源性 TM2 半胱氨酸被改变为甘氨酸的双突变体 C101G/G158C 没有改变 2-APB 激活的动力学。我们认为,在 TM3 位置 158 处引入的半胱氨酸和 TM2 位置 101 处的内源性半胱氨酸之间形成的二硫键阻碍了 Orai3 开放和关闭状态之间的转变。我们的数据提供了这两个残基接近的功能确认,并表明在 Orai3 蛋白内存在对 2-APB 作用敏感的位置。