Lechtreck Karl-Ferdinand, Gould Travis J, Witman George B
Department of Cellular Biology, University of Georgia, 1000 Cedar Ave, 635 Biological Science Bldg,, 30602 Athens, GA, USA.
Cilia. 2013 Nov 27;2(1):15. doi: 10.1186/2046-2530-2-15.
Most motile cilia and flagella have nine outer doublet and two central pair (CP) microtubules. Outer doublet microtubules are continuous with the triplet microtubules of the basal body, are templated by the basal body microtubules, and grow by addition of new subunits to their distal ("plus") ends. In contrast, CP microtubules are not continuous with basal body microtubules, raising the question of how these microtubules are assembled and how their polarity is established.
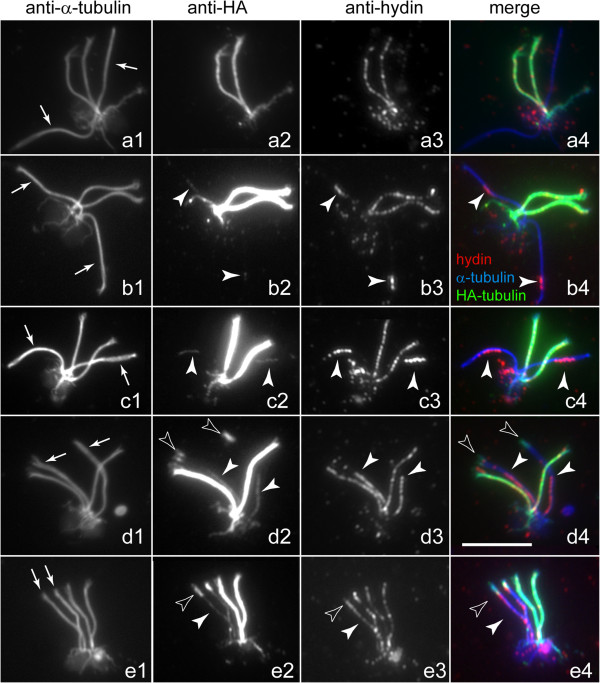
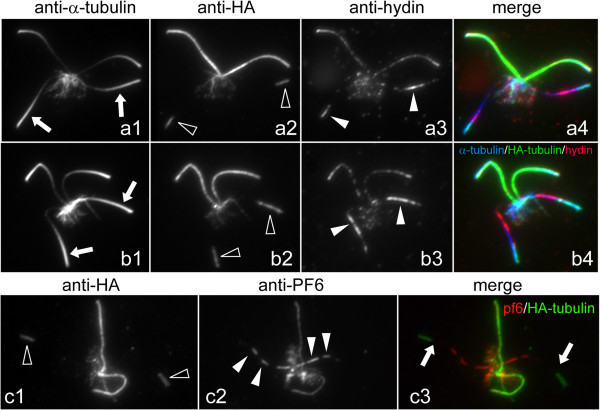
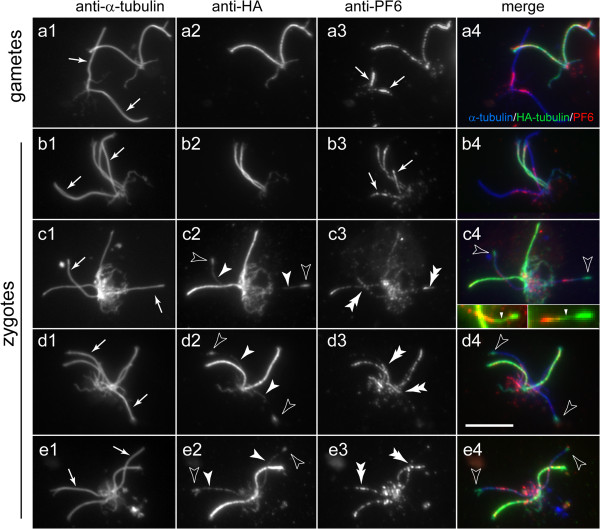
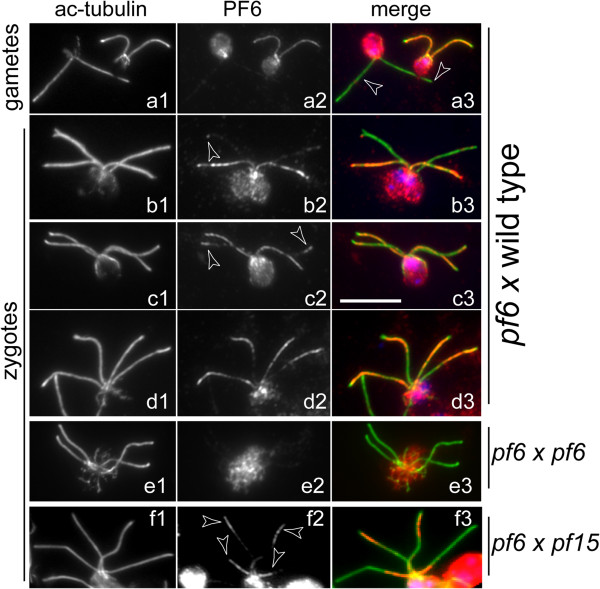
CP assembly in Chlamydomonas reinhardtii was analyzed by electron microscopy and wide-field and super-resolution immunofluorescence microscopy. To analyze CP assembly independently from flagellar assembly, the CP-deficient katanin mutants pf15 or pf19 were mated to wild-type cells. HA-tagged tubulin and the CP-specific protein hydin were used as markers to analyze de novo CP assembly inside the formerly mutant flagella.
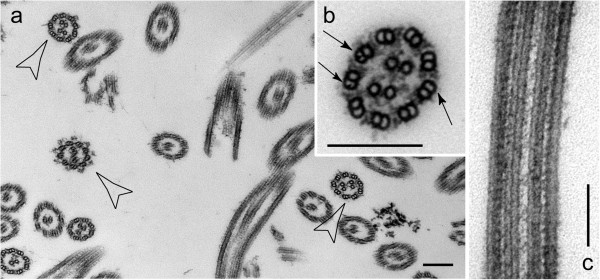
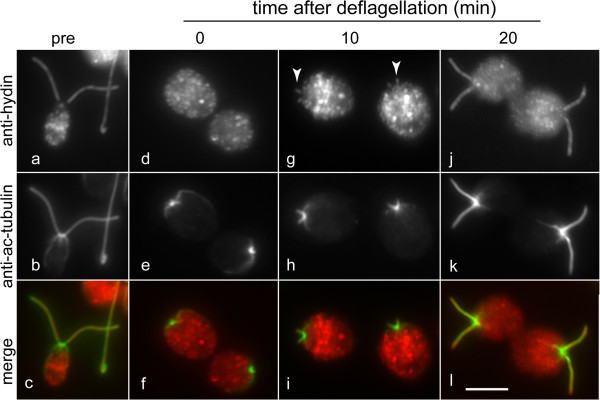
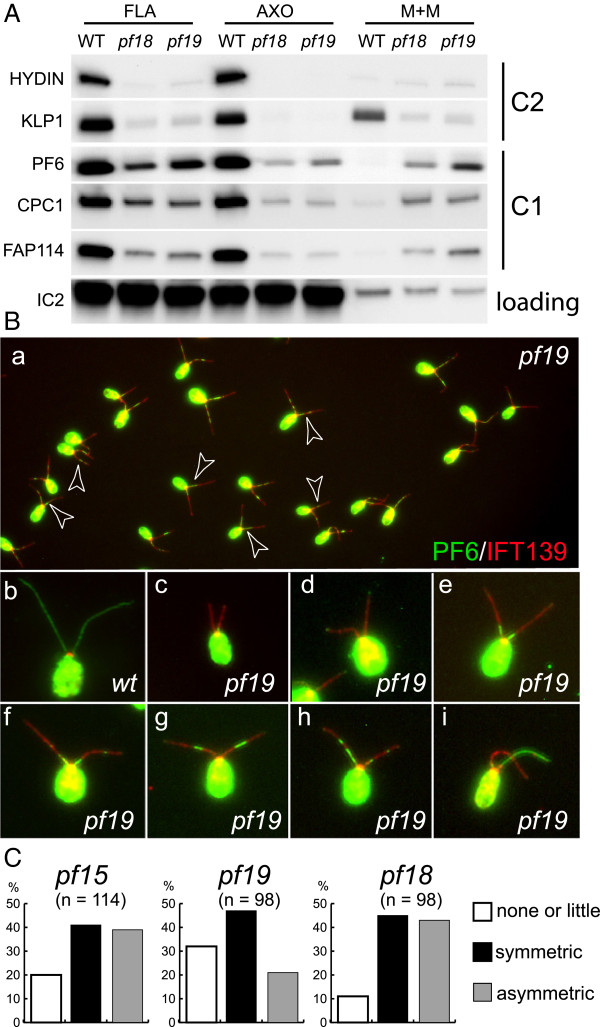
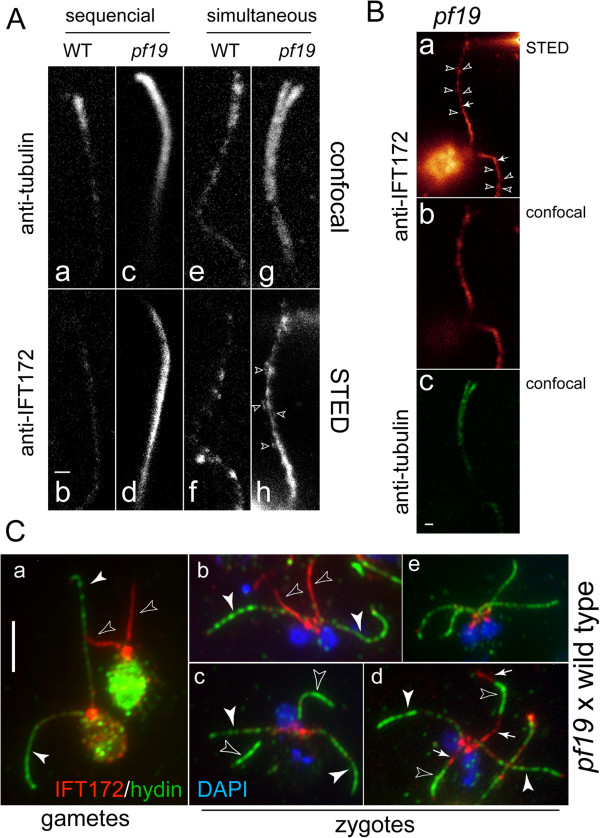
In regenerating flagella, the CP and its projections assemble near the transition zone soon after the onset of outer doublet elongation. During de novo CP assembly in full-length flagella, the nascent CP was first apparent in a subdistal region of the flagellum. The developing CP replaces a fibrous core that fills the axonemal lumen of CP-deficient flagella. The fibrous core contains proteins normally associated with the C1 CP microtubule and proteins involved in intraflagellar transport (IFT). In flagella of the radial spoke-deficient mutant pf14, two pairs of CPs are frequently present with identical correct polarities.
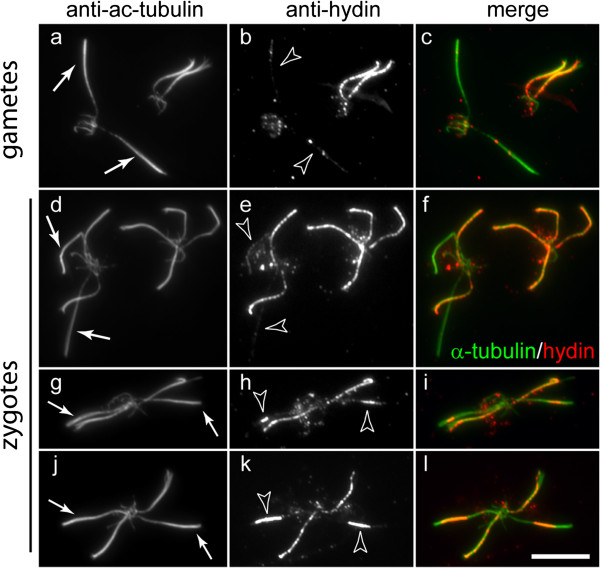
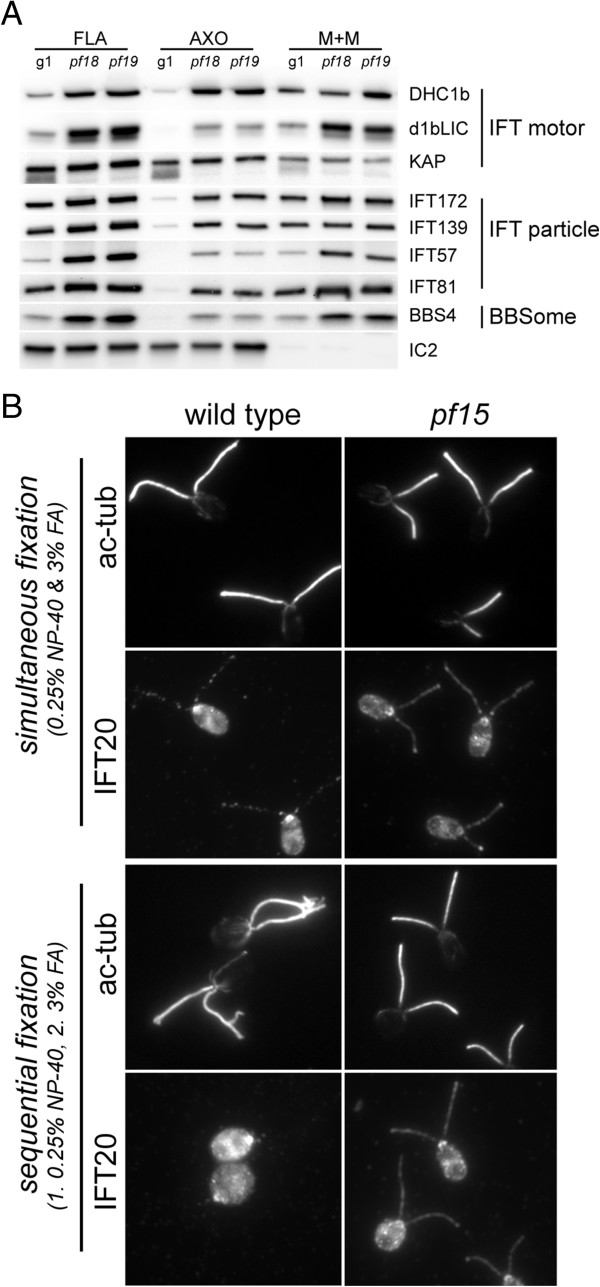
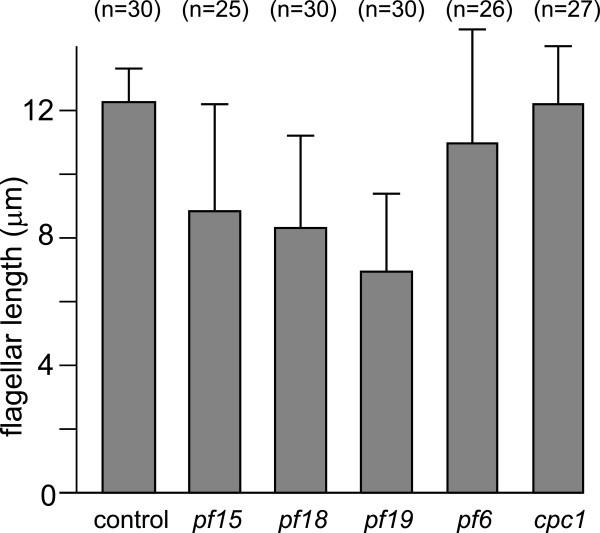
The temporal separation of flagellar and CP assembly in dikaryons formed by mating CP-deficient gametes to wild-type gametes revealed that the formation of the CP does not require proximity to the basal body or transition zone, or to the flagellar tip. The observations on pf14 provide further support that the CP self-assembles without a template and eliminate the possibility that CP polarity is established by interaction with axonemal radial spokes. Polarity of the developing CP may be determined by the proximal-to-distal gradient of precursor molecules. IFT proteins accumulate in flagella of CP mutants; the abnormal distribution of IFT proteins may explain why these flagella are often shorter than normal.
大多数能动纤毛和鞭毛具有九条外周双联微管和两条中央微管对(CP)。外周双联微管与基体的三联微管相连,由基体微管作为模板,并通过在其远端(“正端”)添加新亚基而生长。相比之下,CP微管与基体微管不相连,这就引发了这些微管如何组装以及其极性如何确立的问题。
通过电子显微镜以及宽视野和超分辨率免疫荧光显微镜分析莱茵衣藻中的CP组装。为了独立于鞭毛组装来分析CP组装,将缺乏CP的katanin突变体pf15或pf19与野生型细胞进行交配。带有HA标签的微管蛋白和CP特异性蛋白hydin用作标记物,以分析先前突变鞭毛内的从头CP组装。
在再生鞭毛中,CP及其突起在外周双联微管伸长开始后不久在过渡区附近组装。在全长鞭毛的从头CP组装过程中,新生CP首先在鞭毛的亚远端区域明显可见。发育中的CP取代了填充缺乏CP的鞭毛轴丝腔的纤维状核心。该纤维状核心包含通常与C1 CP微管相关的蛋白质以及参与鞭毛内运输(IFT)的蛋白质。在缺乏径向辐条的突变体pf14的鞭毛中,经常存在两对具有相同正确极性的CP。
通过将缺乏CP的配子与野生型配子交配形成的双核体中鞭毛和CP组装的时间分离表明,CP的形成不需要靠近基体或过渡区,也不需要靠近鞭毛尖端。对pf14突变体的观察结果进一步支持了CP在没有模板的情况下自我组装的观点,并排除了CP极性通过与轴丝径向辐条相互作用而确立的可能性。发育中CP的极性可能由前体分子的近端到远端梯度决定。IFT蛋白在CP突变体的鞭毛中积累;IFT蛋白的异常分布可能解释了为什么这些鞭毛通常比正常鞭毛短。