Li Stephen Yt, Mruk Dolores D, Cheng C Yan
The Mary M. Wohlford Laboratory for Male Contraceptive Research; Center for Biomedical Research; Population Council; New York, NY USA.
Spermatogenesis. 2013 Jul 1;3(3):e25385. doi: 10.4161/spmg.25385. Epub 2013 Jun 21.
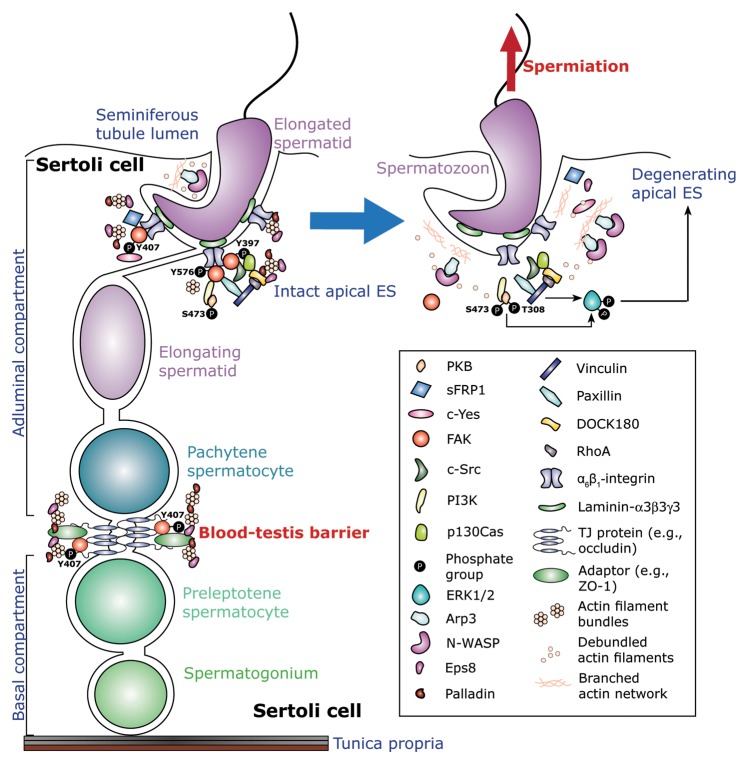
During spermatogenesis, spermatogonia (2n, diploid) undergo a series of mitotic divisions as well as differentiation to become spermatocytes, which enter meiosis I to be followed by meiosis II to form round spermatids (1n, haploid), and then differentiate into spermatozoa (1n, haploid) via spermiogenesis. These events take place in the epithelium of the seminiferous tubule, involving extensive junction restructuring at the Sertoli-Sertoli and Sertoli-germ cell interface to allow the transport of developing germ cells across the epithelium. Although structural aspects of these cell-cell junctions have been studied, the underlying mechanism(s) that governs these events has yet to be explored. Earlier studies have shown that a non-receptor protein tyrosine kinase known as focal adhesion kinase (FAK) is a likely regulator of these events due to the stage-specific and spatiotemporal expression of its various phosphorylated/activated forms at the testis-specific anchoring junctions in the testis, as well as its association with actin regulatory proteins. Recent studies have shown that FAK, in particular its two activated phosphorylated forms p-FAK-Tyr and p-FAK-Tyr, are crucial regulators in modulating junction restructuring at the Sertoli cell-cell interface at the blood-testis barrier (BTB) known as the basal ectoplasmic specialization (basal ES), as well as at the Sertoli-spermatid interface called apical ES during spermiogenesis via its effects on the filamentous (F)-actin organization at the ES. We herein summarize and critically evaluate the current knowledge regarding the physiological significance of FAK in regulating BTB and apical ES dynamics by governing the conversion of actin filaments at the ES from a "bundled" to a "de-bundled/branched" configuration and vice versa. We also provide a molecular model on the role of FAK in regulating these events based on the latest findings in the field.
在精子发生过程中,精原细胞(2n,二倍体)经历一系列有丝分裂以及分化,成为精母细胞,精母细胞进入减数分裂I,随后进行减数分裂II,形成圆形精子细胞(1n,单倍体),然后通过精子形成过程分化为精子(1n,单倍体)。这些事件发生在生精小管的上皮中,涉及支持细胞-支持细胞和支持细胞-生殖细胞界面处广泛的连接重组,以允许发育中的生殖细胞穿过上皮运输。尽管已经研究了这些细胞间连接的结构方面,但控制这些事件的潜在机制尚未得到探索。早期研究表明,一种称为粘着斑激酶(FAK)的非受体蛋白酪氨酸激酶可能是这些事件的调节因子,这是由于其各种磷酸化/激活形式在睾丸中睾丸特异性锚定连接处的阶段特异性和时空表达,以及它与肌动蛋白调节蛋白的关联。最近的研究表明,FAK,特别是其两种激活的磷酸化形式p-FAK-Tyr和p-FAK-Tyr,是调节血睾屏障(BTB)处支持细胞-细胞界面(称为基底外质特化,basal ES)以及精子形成过程中支持细胞-精子细胞界面(称为顶端ES)处连接重组的关键调节因子,它通过影响ES处的丝状(F)-肌动蛋白组织来实现。我们在此总结并批判性地评估了关于FAK通过控制ES处肌动蛋白丝从“束状”到“解束/分支”构型的转变以及反之亦然来调节BTB和顶端ES动态的生理意义的当前知识。我们还根据该领域的最新发现提供了一个关于FAK在调节这些事件中作用的分子模型。