Lazarus David, Barron John, Renaudie Johan, Diver Patrick, Türke Andreas
Museum für Naturkunde, Berlin, Germany.
United States Geological Survey, Menlo Park, California, United States of America.
PLoS One. 2014 Jan 22;9(1):e84857. doi: 10.1371/journal.pone.0084857. eCollection 2014.
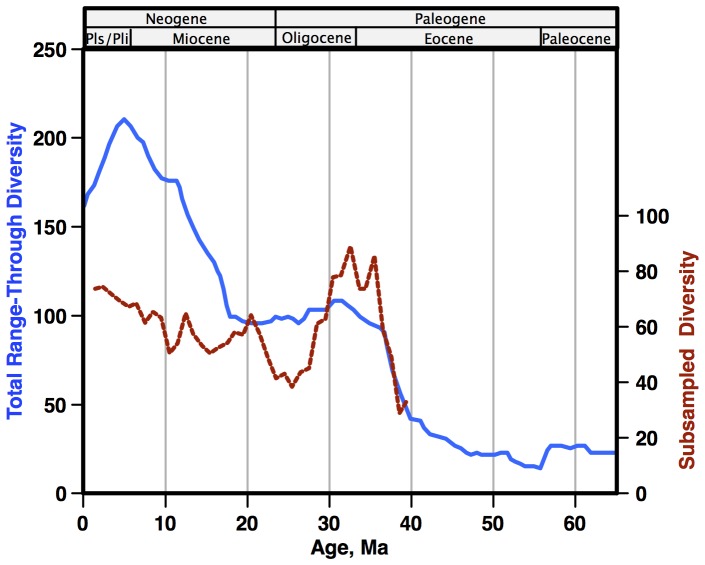
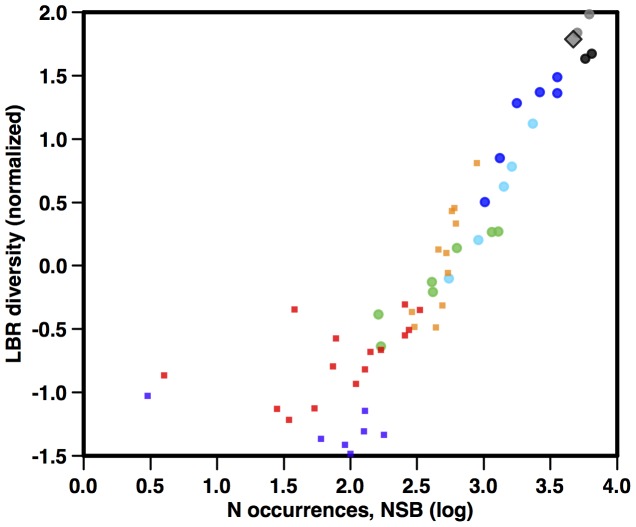
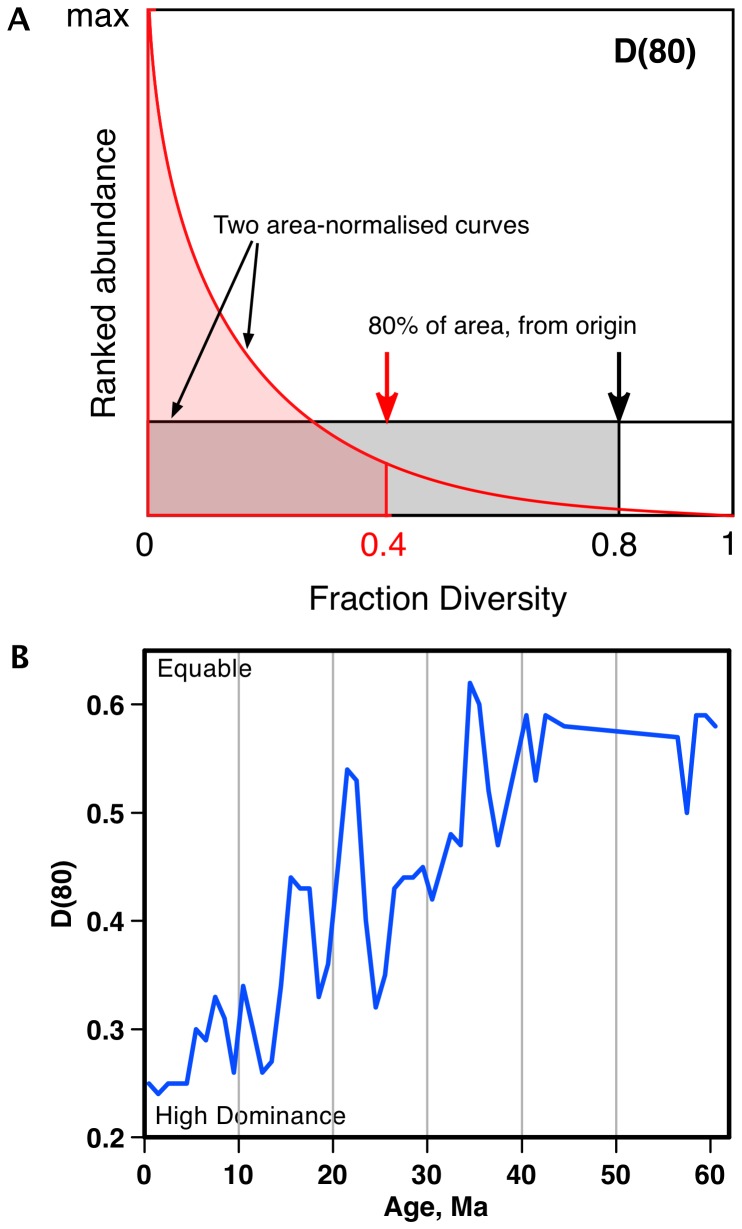
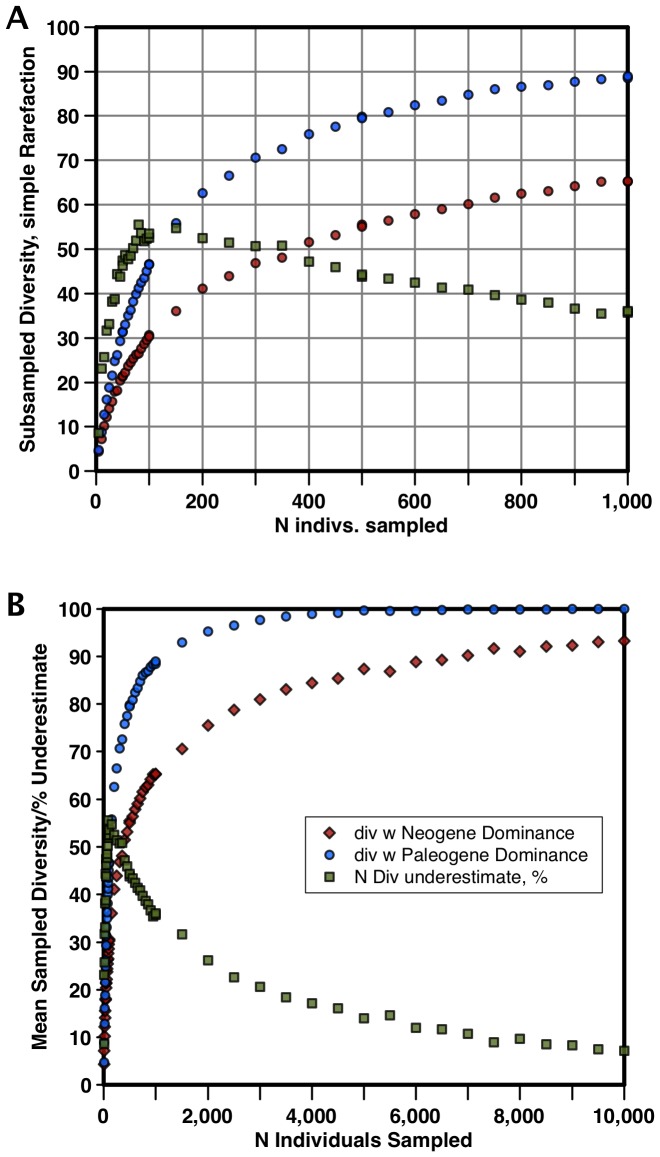
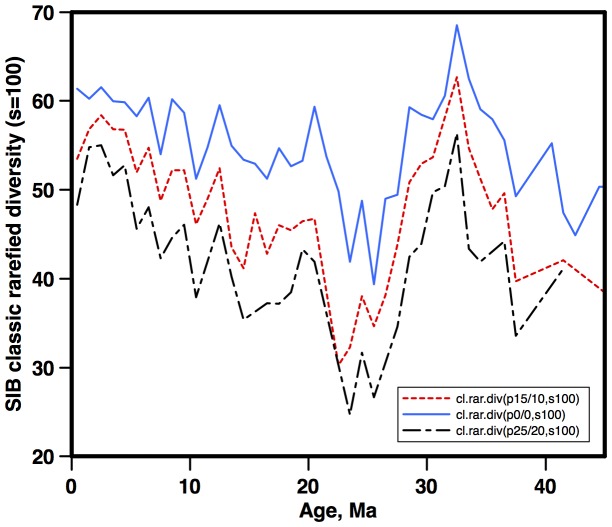
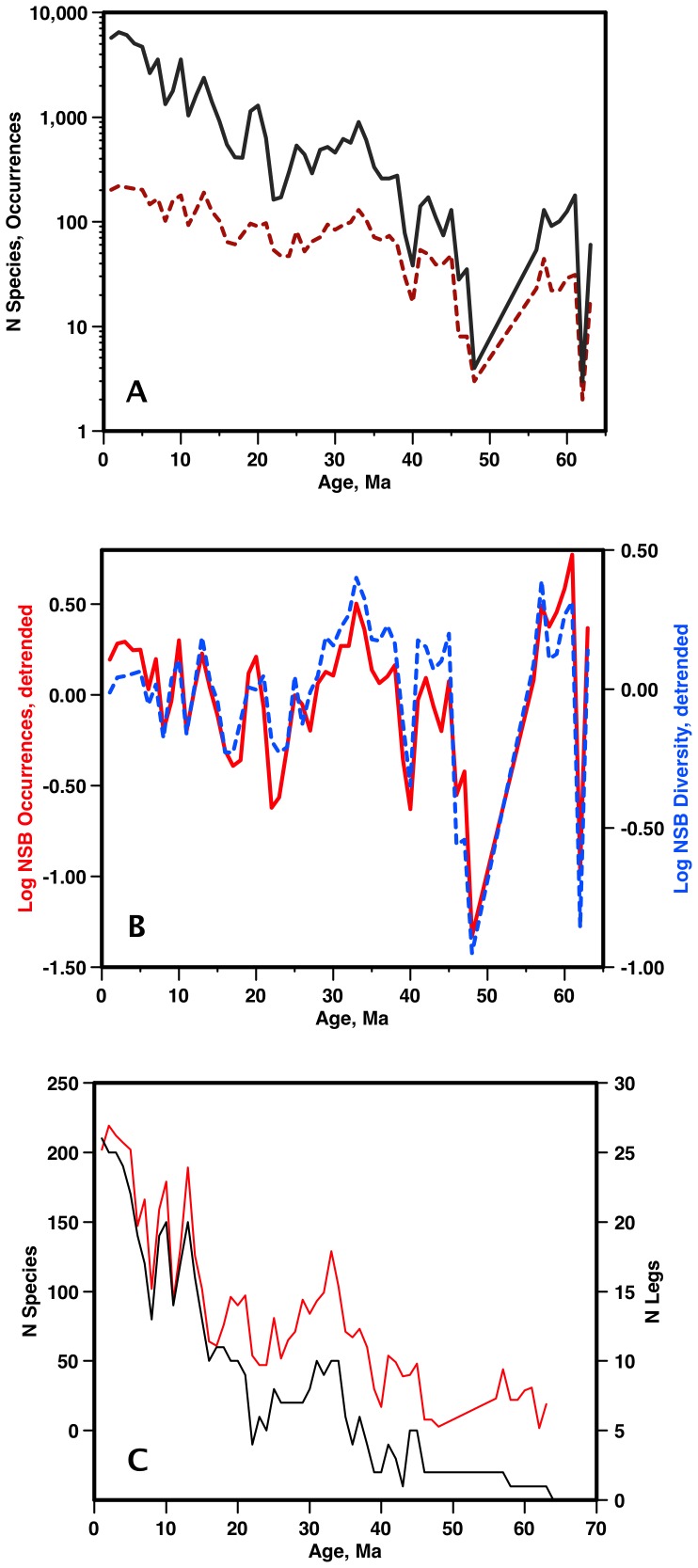
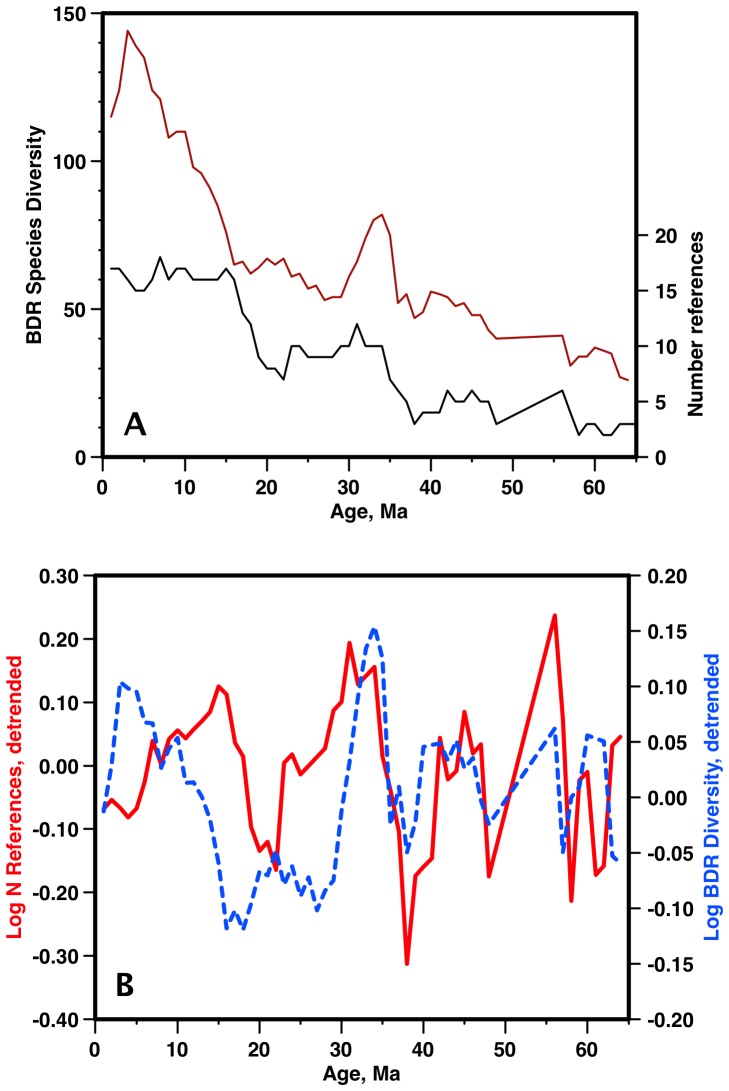
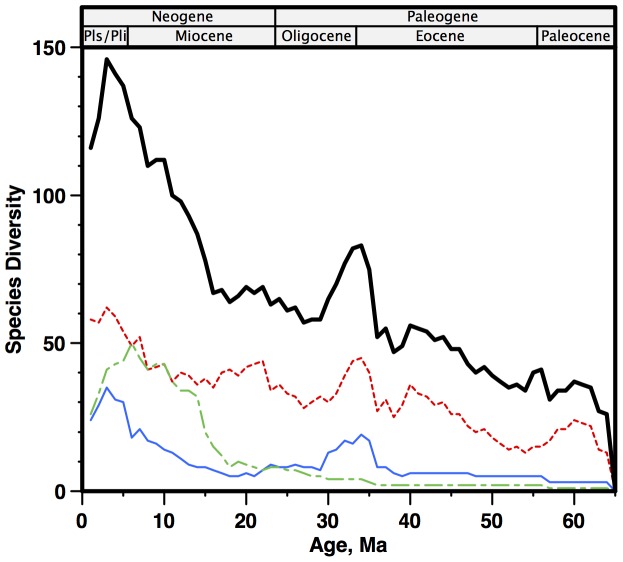
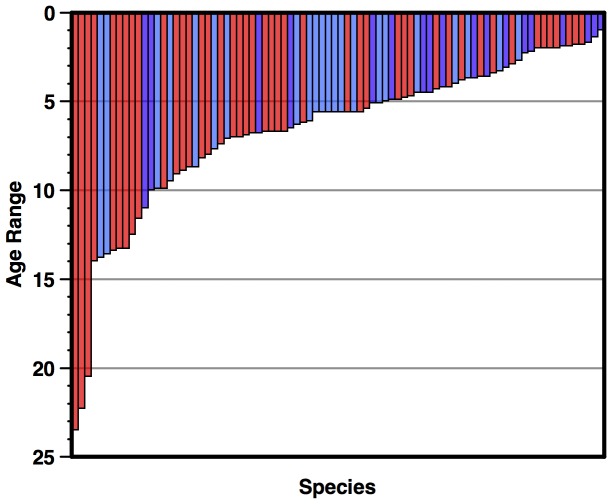
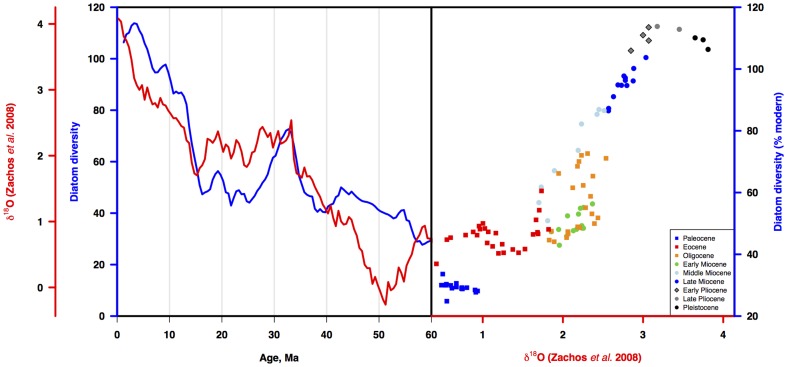
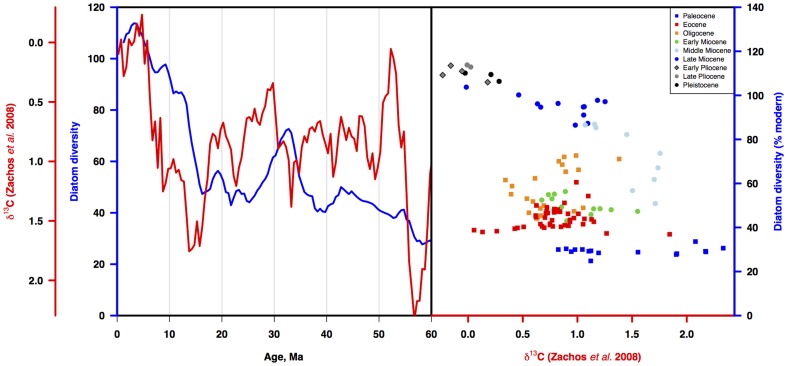
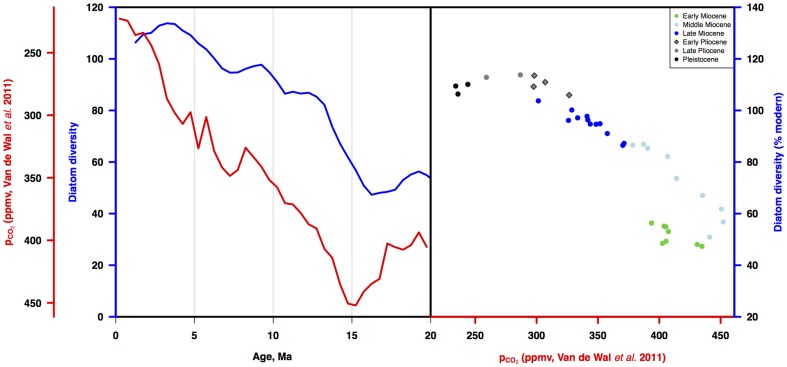
Marine planktonic diatoms export carbon to the deep ocean, playing a key role in the global carbon cycle. Although commonly thought to have diversified over the Cenozoic as global oceans cooled, only two conflicting quantitative reconstructions exist, both from the Neptune deep-sea microfossil occurrences database. Total diversity shows Cenozoic increase but is sample size biased; conventional subsampling shows little net change. We calculate diversity from a separately compiled new diatom species range catalog, and recalculate Neptune subsampled-in-bin diversity using new methods to correct for increasing Cenozoic geographic endemism and decreasing Cenozoic evenness. We find coherent, substantial Cenozoic diversification in both datasets. Many living cold water species, including species important for export productivity, originate only in the latest Miocene or younger. We make a first quantitative comparison of diatom diversity to the global Cenozoic benthic ∂(18)O (climate) and carbon cycle records (∂(13)C, and 20-0 Ma pCO2). Warmer climates are strongly correlated with lower diatom diversity (raw: rho = .92, p<.001; detrended, r = .6, p = .01). Diatoms were 20% less diverse in the early late Miocene, when temperatures and pCO2 were only moderately higher than today. Diversity is strongly correlated to both ∂(13)C and pCO2 over the last 15 my (for both: r>.9, detrended r>.6, all p<.001), but only weakly over the earlier Cenozoic, suggesting increasingly strong linkage of diatom and climate evolution in the Neogene. Our results suggest that many living marine planktonic diatom species may be at risk of extinction in future warm oceans, with an unknown but potentially substantial negative impact on the ocean biologic pump and oceanic carbon sequestration. We cannot however extrapolate our my-scale correlations with generic climate proxies to anthropogenic time-scales of warming without additional species-specific information on proximate ecologic controls.
海洋浮游硅藻将碳输送到深海,在全球碳循环中发挥着关键作用。尽管通常认为随着全球海洋变冷,硅藻在新生代实现了多样化,但目前仅有两个相互矛盾的定量重建结果,且均来自海王星深海微化石出现数据库。总多样性显示新生代有所增加,但存在样本量偏差;传统的二次抽样显示净变化不大。我们根据单独编制的新硅藻物种分布目录计算多样性,并使用新方法重新计算海王星按箱二次抽样的多样性,以校正新生代地理特有性增加和新生代均匀度降低的情况。我们在两个数据集中都发现了一致且显著的新生代多样化现象。许多现存的冷水物种,包括对输出生产力很重要的物种,直到最新的中新世或更晚才出现。我们首次对硅藻多样性与全球新生代底栖δ(18)O(气候)和碳循环记录(δ(13)C以及20 - 0 Ma的pCO2)进行了定量比较。温暖的气候与较低的硅藻多样性密切相关(原始数据:rho = 0.92,p < 0.001;去趋势后,r = 0.6,p = 0.01)。在中新世晚期早期,当温度和pCO2仅略高于现今时,硅藻的多样性降低了20%。在过去1500万年中,多样性与δ(13)C和pCO2都密切相关(两者:r > 0.9,去趋势后r > 0.6,所有p < 0.001),但在新生代早期相关性较弱,这表明在新近纪硅藻与气候演化的联系越来越紧密。我们的结果表明,在未来温暖的海洋中,许多现存的海洋浮游硅藻物种可能面临灭绝风险,这对海洋生物泵和海洋碳固存将产生未知但可能很大的负面影响。然而,在没有关于近期生态控制的额外物种特异性信息的情况下,我们无法将我们在百万年级别上与通用气候指标的相关性外推到人为变暖的时间尺度上。