Watson School of Biological Sciences.
Genes Dev. 2014 Apr 1;28(7):797-807. doi: 10.1101/gad.235622.113.
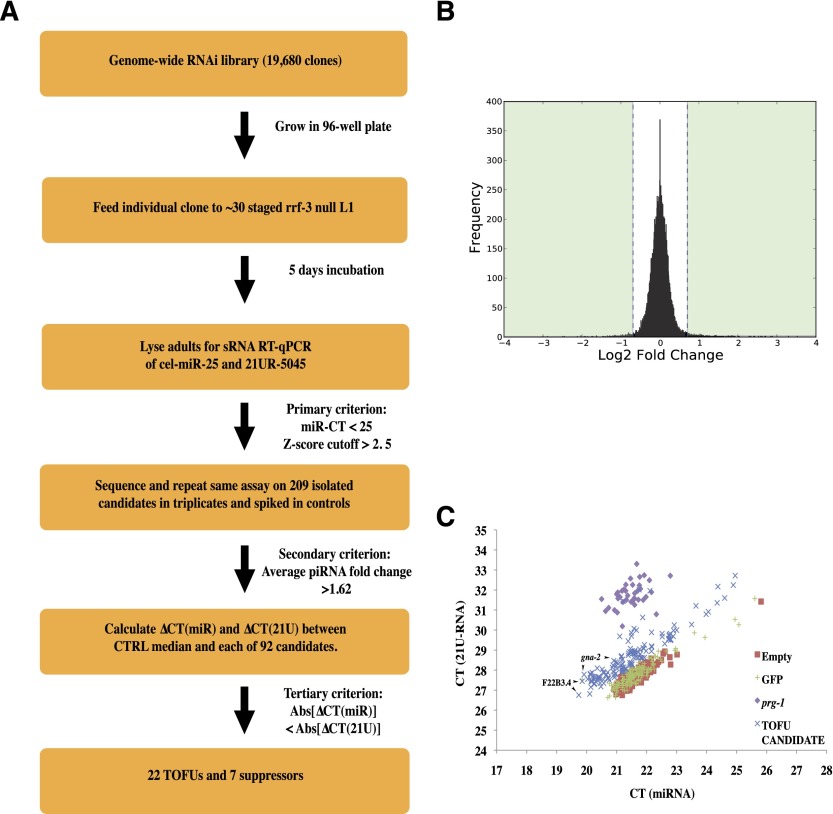
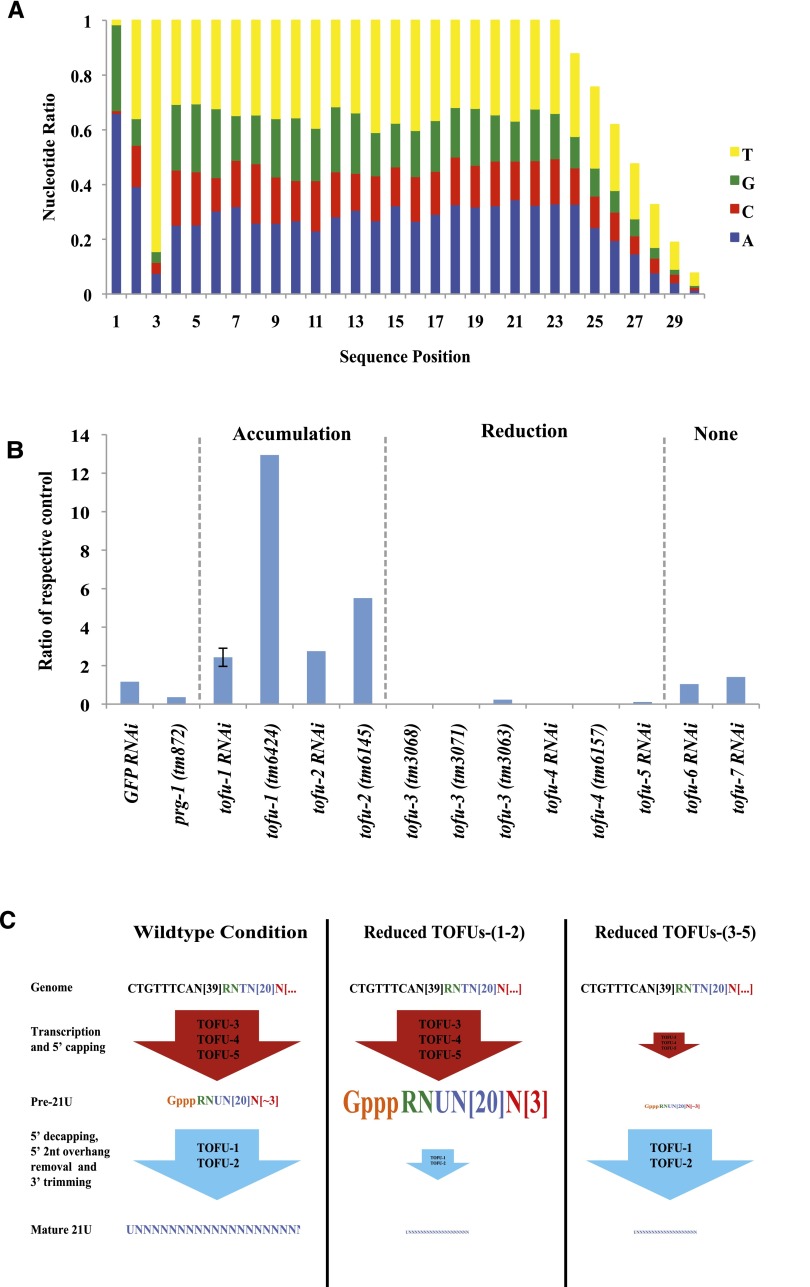
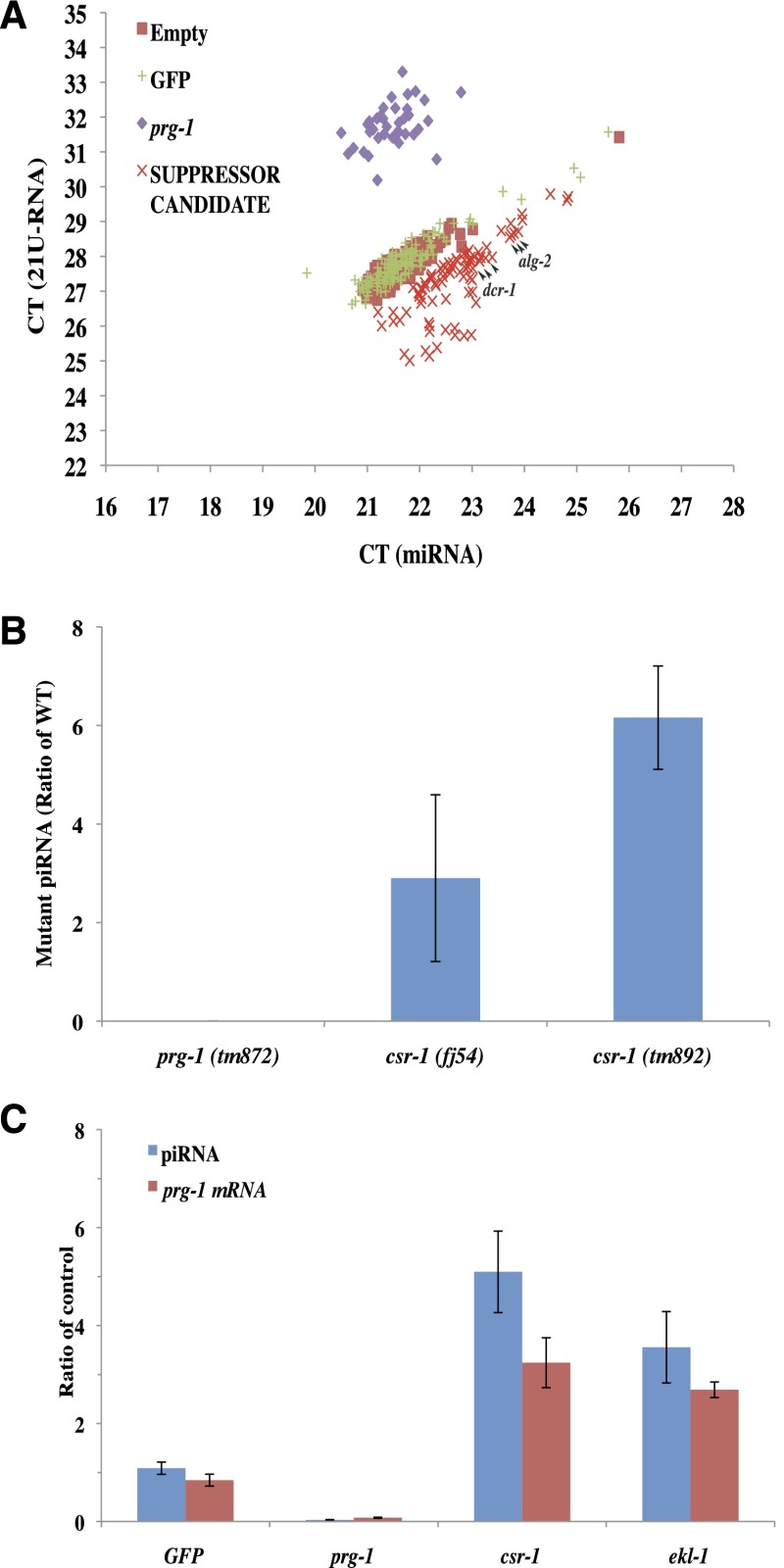
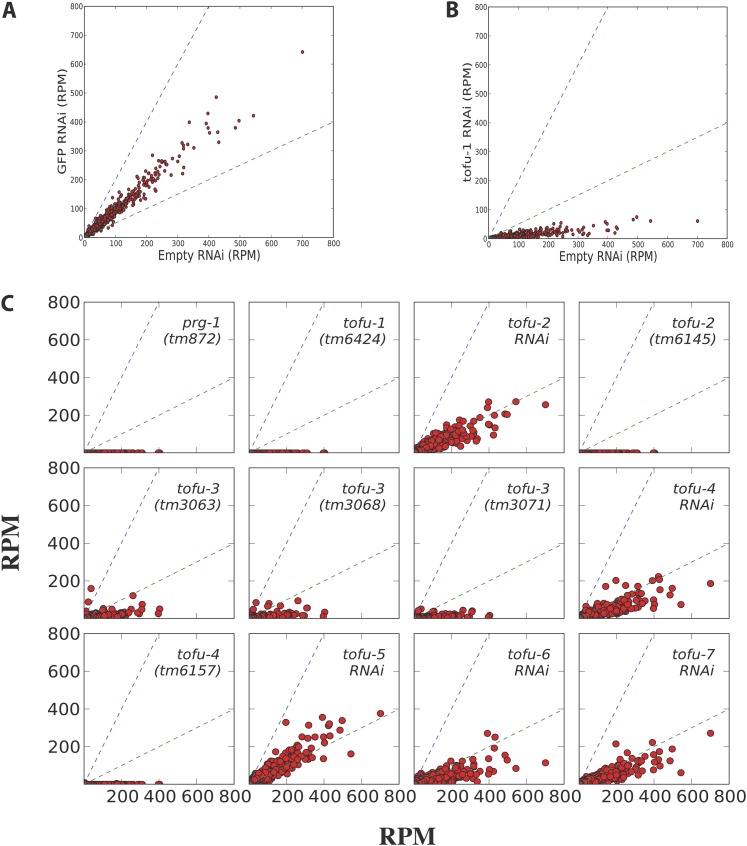
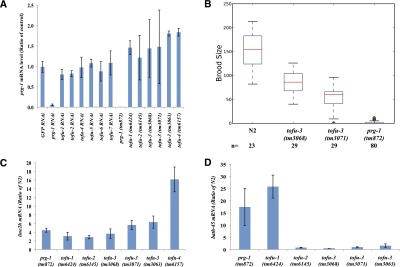
In animals, piRNAs and their associated Piwi proteins guard germ cell genomes against mobile genetic elements via an RNAi-like mechanism. In Caenorhabditis elegans, 21U-RNAs comprise the piRNA class, and these collaborate with 22G RNAs via unclear mechanisms to discriminate self from nonself and selectively and heritably silence the latter. Recent work indicates that 21U-RNAs are post-transcriptional processing products of individual transcription units that produce ∼ 26-nucleotide capped precursors. However, nothing is known of how the expression of precursors is controlled or how primary transcripts give rise to mature small RNAs. We conducted a genome-wide RNAi screen to identify components of the 21U biogenesis machinery. Screening by direct, quantitative PCR (qPCR)-based measurements of mature 21U-RNA levels, we identified 22 genes important for 21U-RNA production, termed TOFUs (Twenty-One-u Fouled Ups). We also identified seven genes that normally repress 21U production. By measuring mature 21U-RNA and precursor levels for the seven strongest hits from the screen, we assigned factors to discrete stages of 21U-RNA production. Our work identifies for the first time factors separately required for the transcription of 21U precursors and the processing of these precursors into mature 21U-RNAs, thereby providing a resource for studying the biogenesis of this important small RNA class.
在动物中,piRNA 及其相关的 Piwi 蛋白通过类似于 RNAi 的机制保护生殖细胞基因组免受移动遗传元件的侵害。在秀丽隐杆线虫中,21U-RNAs 构成了 piRNA 类,这些 RNA 与 22G RNA 合作,通过不清楚的机制来区分自我和非自我,并选择性地和遗传性地沉默后者。最近的工作表明,21U-RNAs 是单个转录单元产生的约 26 个核苷酸加帽前体的转录后加工产物。然而,人们对前体的表达如何受到控制,以及初级转录本如何产生成熟的小 RNA 一无所知。我们进行了全基因组 RNAi 筛选,以鉴定 21U 生物发生机制的组成部分。通过直接、基于定量 PCR(qPCR)的成熟 21U-RNA 水平的定量测量进行筛选,我们鉴定了 22 个对 21U-RNA 产生重要的基因,称为 TOFUs(Twenty-One-u Fouled Ups)。我们还鉴定了七个通常抑制 21U 产生的基因。通过测量筛选中七个最强命中的成熟 21U-RNA 和前体水平,我们将因子分配到 21U-RNA 产生的离散阶段。我们的工作首次鉴定了分别需要转录 21U 前体和将这些前体加工成成熟 21U-RNAs 的因子,从而为研究这一重要小 RNA 类的生物发生提供了资源。