Schneikert Jean, Ruppert Jan Gustav, Behrens Jürgen, Wenzel Eva Maria
Nikolaus-Fiebiger-Center for Molecular Medicine, University of Erlangen-Nürnberg, Erlangen, Germany.
Centre for Cancer Biomedicine, Faculty of Medicine, University of Oslo, Oslo, Norway.
PLoS One. 2014 Apr 10;9(4):e94413. doi: 10.1371/journal.pone.0094413. eCollection 2014.
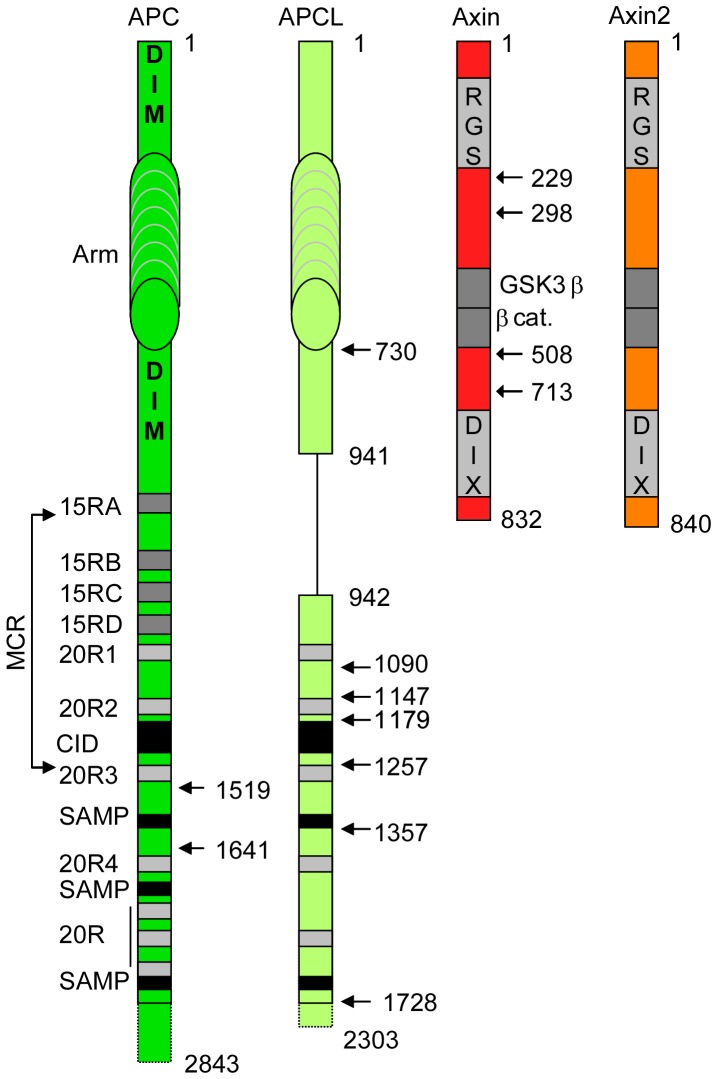
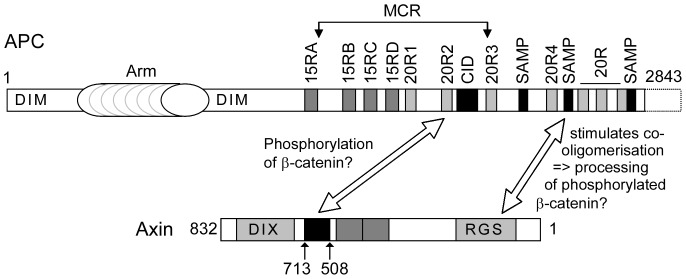
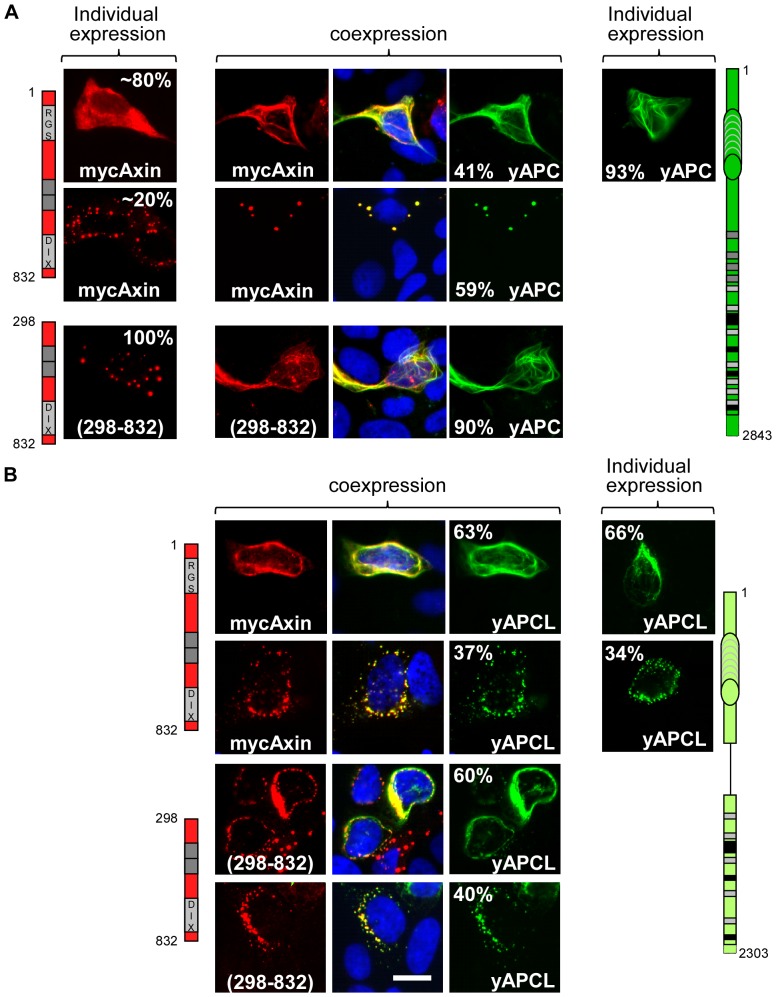
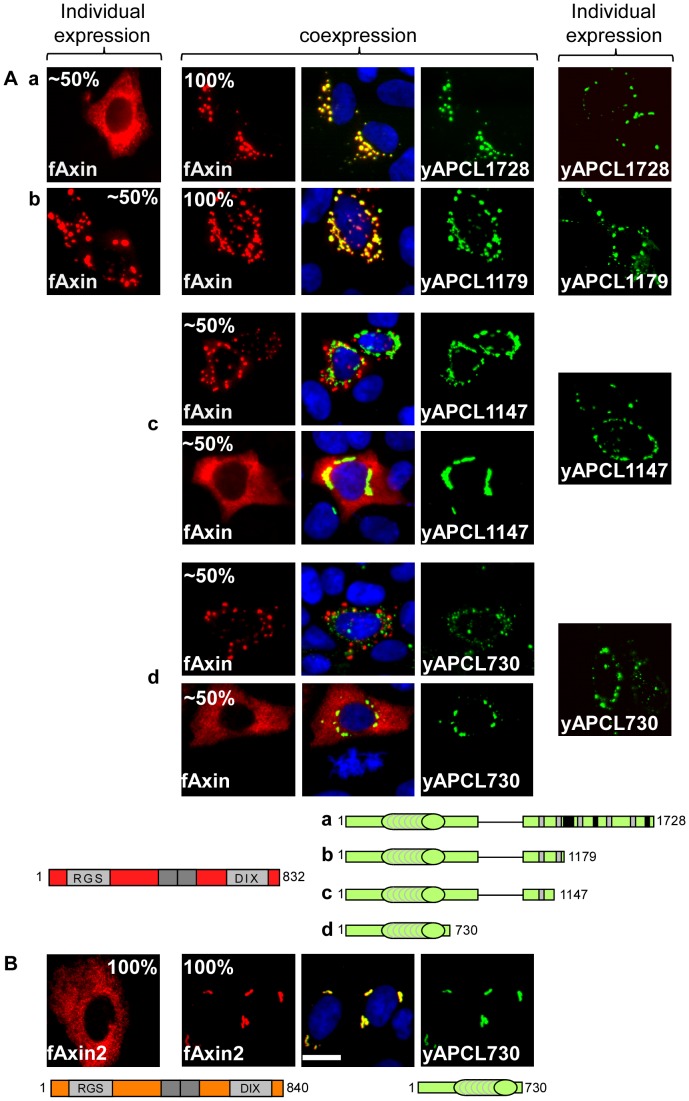
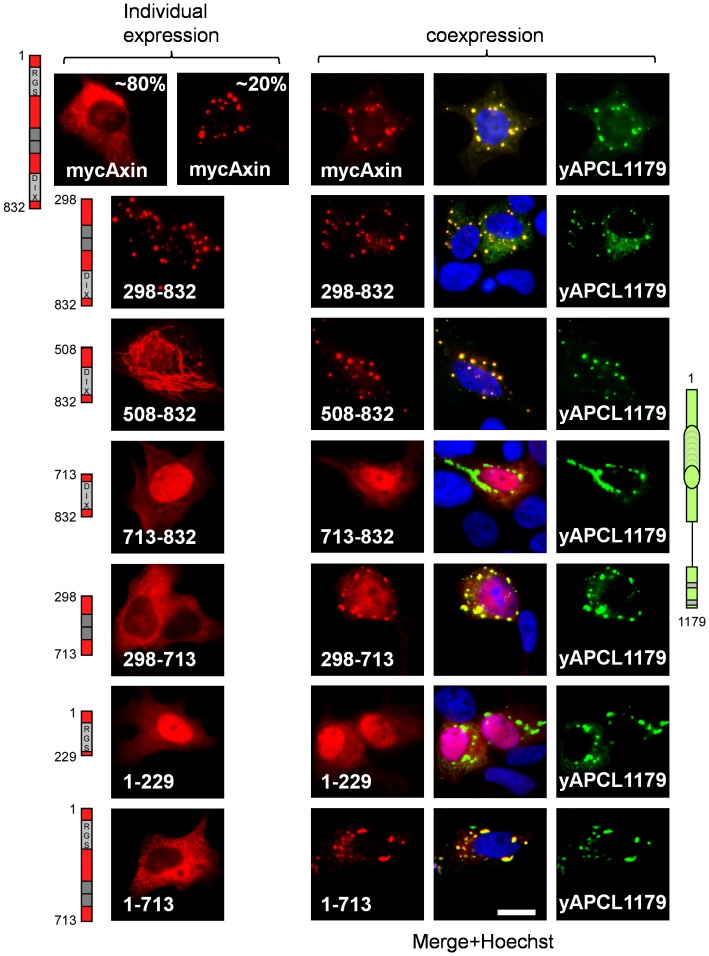
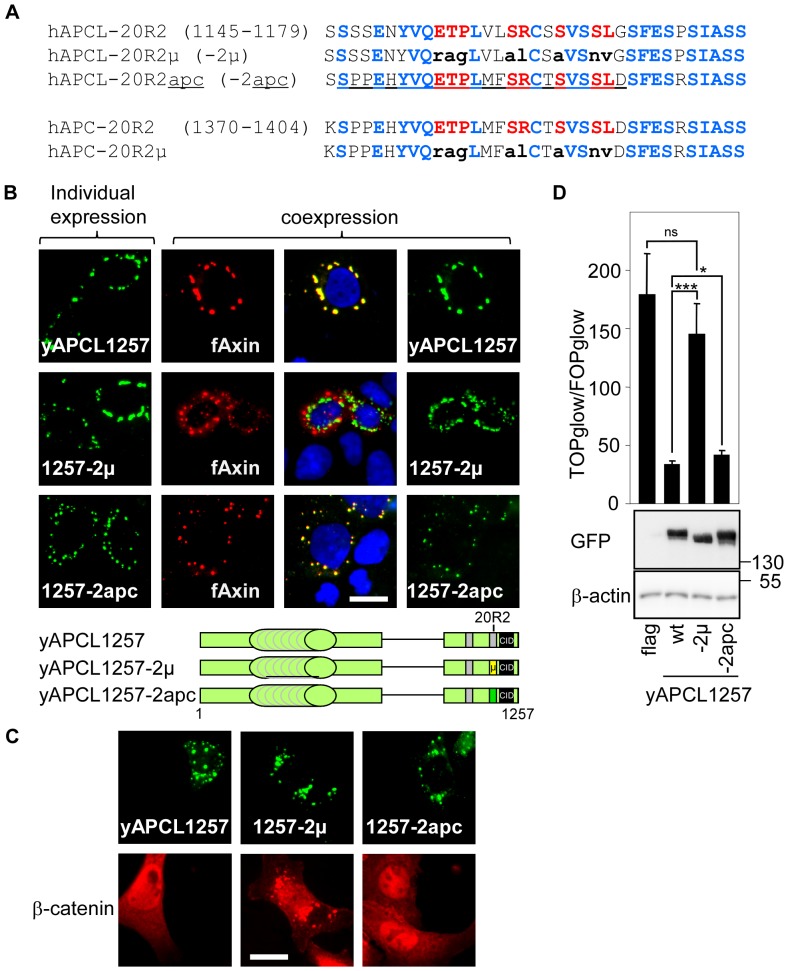
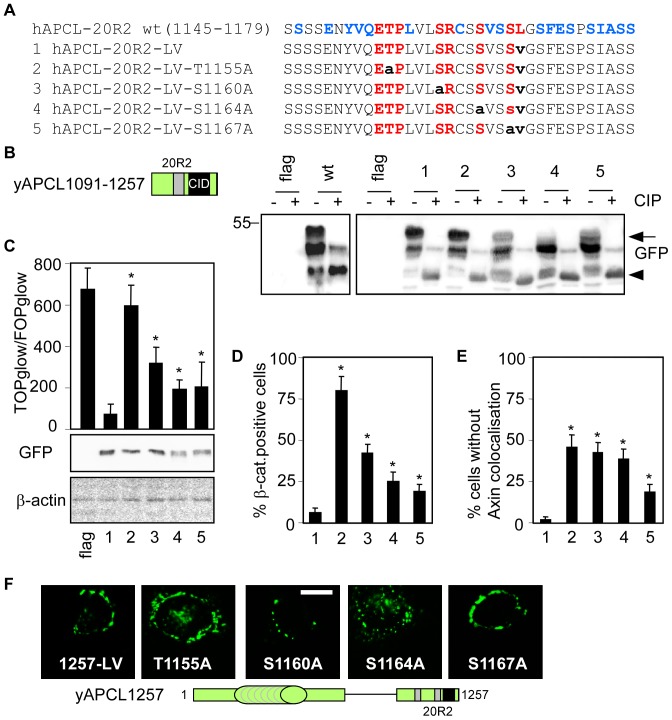
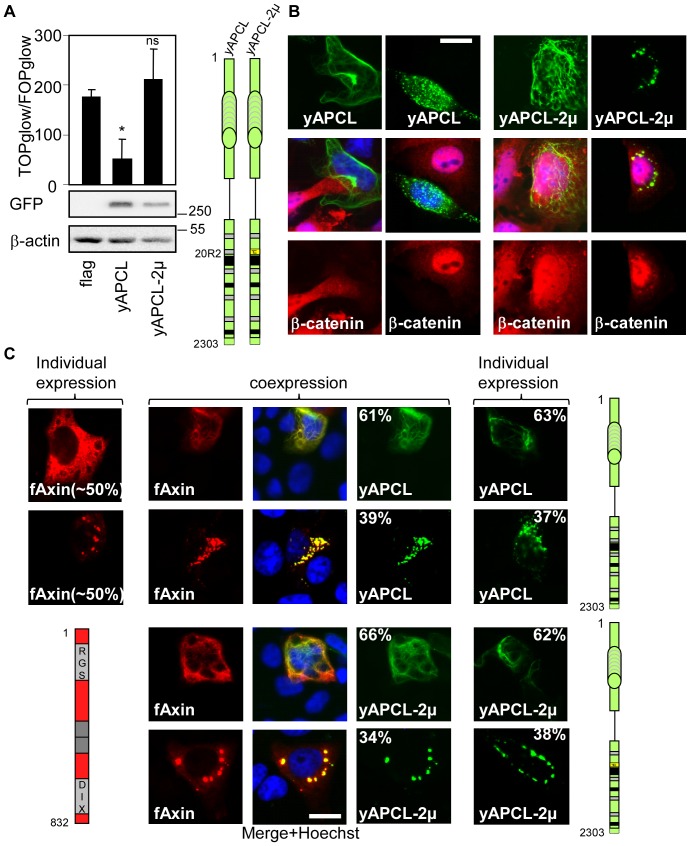
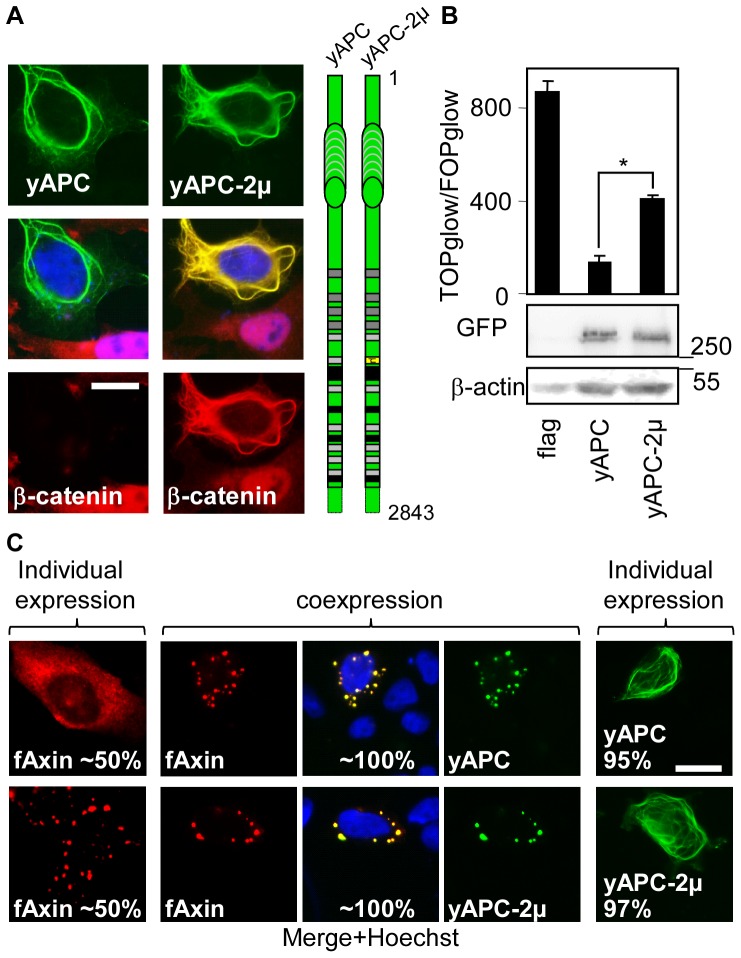
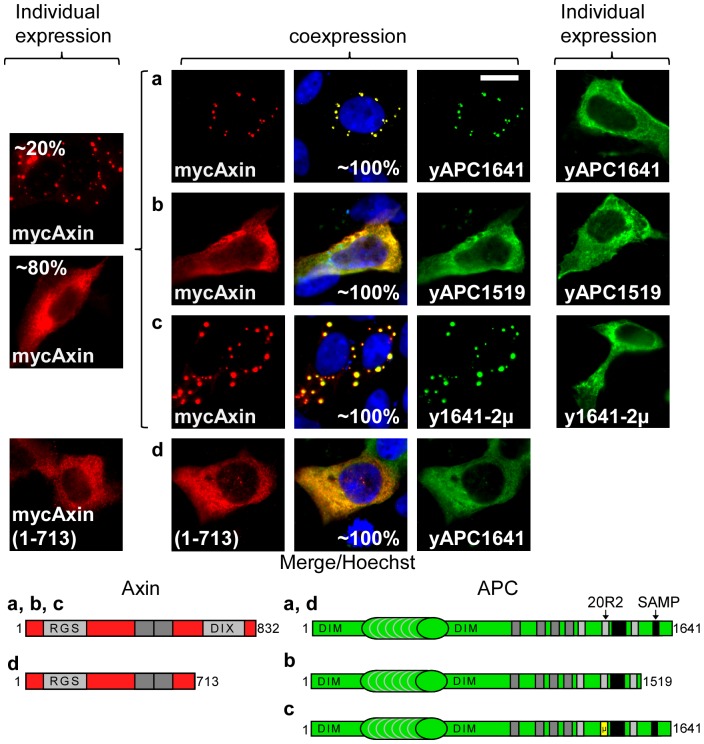
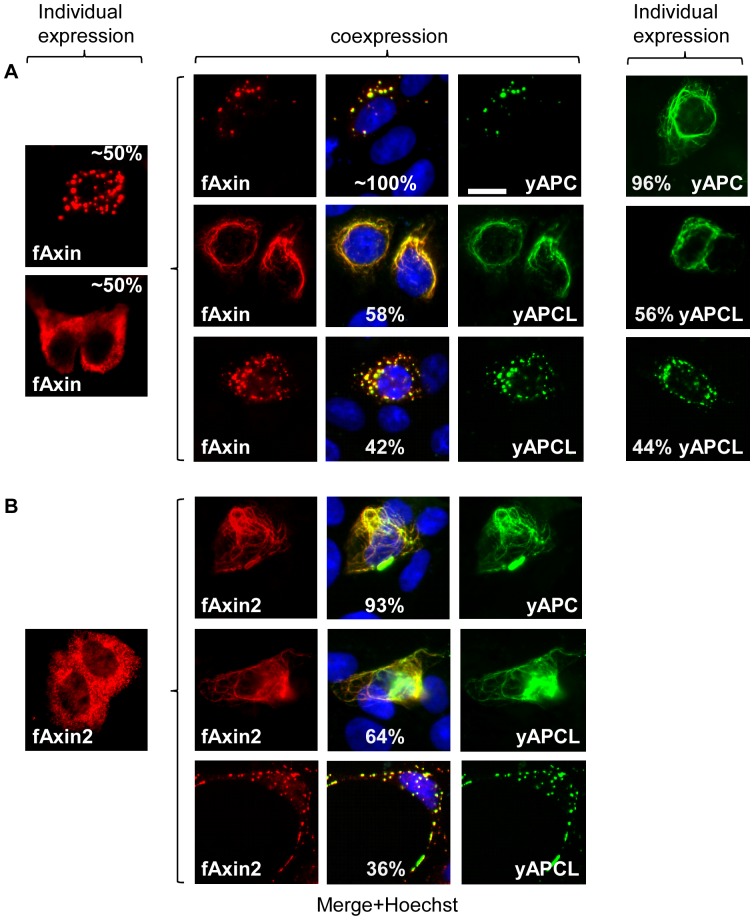
Wnt signalling is prevented by the proteosomal degradation of β-catenin, which occurs in a destruction complex containing adenomatous polyposis coli (APC), APC-like (APCL), Axin and Axin2. Truncating mutations of the APC gene result in the constitutive stabilisation of β-catenin and the initiation of colon cancer, although tumour cells tolerate the expression of wild-type APCL. Using the colocalisation of overexpressed Axin, APC and APCL constructs as a readout of interaction, we found that Axin interacted with the second twenty amino acid repeat (20R2) of APC and APCL. This interaction involved a domain adjacent to the C-terminal DIX domain of Axin. We identified serine residues within the 20R2 of APCL that were involved in Axin colocalisation, the phosphorylation of truncated APCL and the down-regulation of β-catenin. Our results indicated that Axin, but not Axin2, displaced APC, but not APCL, from the cytoskeleton and stimulated its incorporation into bright cytoplasmic dots that others have recognised as β-catenin destruction complexes. The SAMP repeats in APC interact with the N-terminal RGS domain of Axin. Our data showed that a short domain containing the first SAMP repeat in truncated APC was required to stimulate Axin oligomerisation. This was independent of Axin colocalisation with 20R2. Our data also suggested that the RGS domain exerted an internal inhibitory constraint on Axin oligomerisation. Considering our data and those from others, we discuss a working model whereby β-catenin phosphorylation involves Axin and the 20R2 of APC or APCL and further processing of phospho-β-catenin occurs upon the oligomerisation of Axin that is induced by binding the SAMP repeats in APC.
Wnt信号传导通过β-连环蛋白的蛋白酶体降解来阻止,这种降解发生在一个包含腺瘤性息肉病蛋白(APC)、APC样蛋白(APCL)、轴蛋白(Axin)和Axin2的破坏复合物中。APC基因的截短突变导致β-连环蛋白的组成型稳定和结肠癌的起始,尽管肿瘤细胞能够耐受野生型APCL的表达。以过表达的Axin、APC和APCL构建体的共定位作为相互作用的读数,我们发现Axin与APC和APCL的第二个二十氨基酸重复序列(20R2)相互作用。这种相互作用涉及Axin C末端DIX结构域附近的一个结构域。我们确定了APCL的20R2内的丝氨酸残基,这些残基参与Axin共定位、截短的APCL的磷酸化以及β-连环蛋白的下调。我们的结果表明,Axin而非Axin2从细胞骨架上取代了APC而非APCL,并刺激其并入明亮的细胞质点,其他人已将这些点识别为β-连环蛋白破坏复合物。APC中的SAMP重复序列与Axin的N末端RGS结构域相互作用。我们的数据表明,截短的APC中包含第一个SAMP重复序列的短结构域是刺激Axin寡聚化所必需的。这与Axin与20R2的共定位无关。我们的数据还表明,RGS结构域对Axin寡聚化施加了内部抑制性限制。考虑到我们的数据和其他人的数据,我们讨论了一个工作模型,即β-连环蛋白磷酸化涉及Axin和APC或APCL的20R2,并且磷酸化β-连环蛋白的进一步加工发生在由APC中的SAMP重复序列结合诱导的Axin寡聚化时。