Sampathkumar Arun, Krupinski Pawel, Wightman Raymond, Milani Pascale, Berquand Alexandre, Boudaoud Arezki, Hamant Olivier, Jönsson Henrik, Meyerowitz Elliot M
Division of Biology and Biological Engineering, California Institute of Technology, Pasadena, United States.
Elife. 2014 Apr 16;3:e01967. doi: 10.7554/eLife.01967.
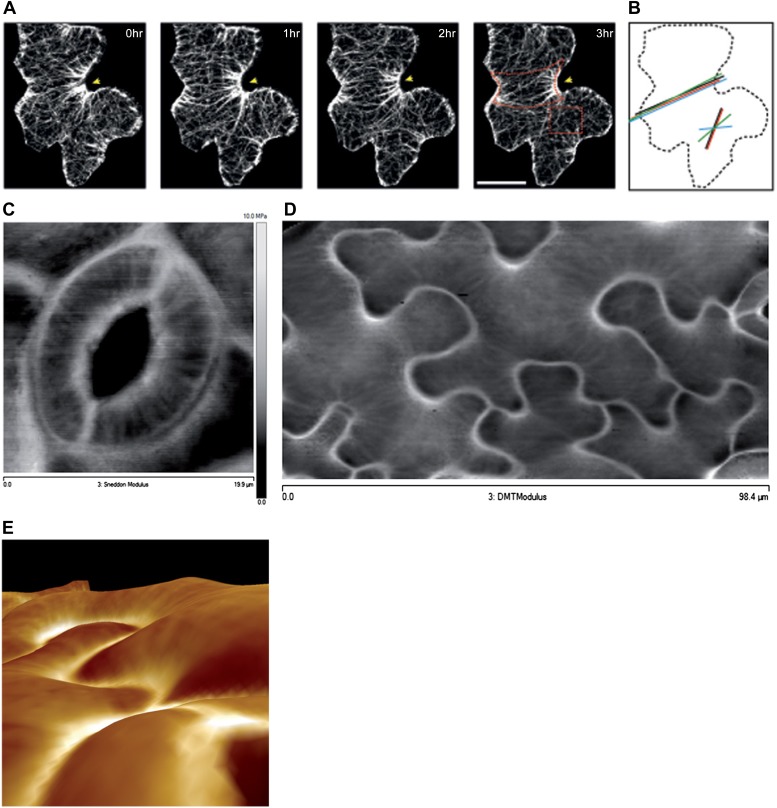
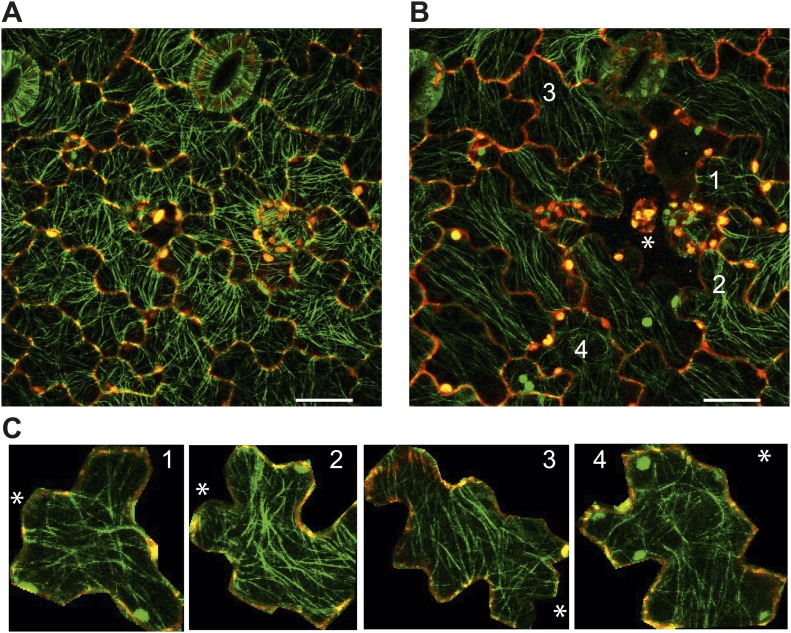
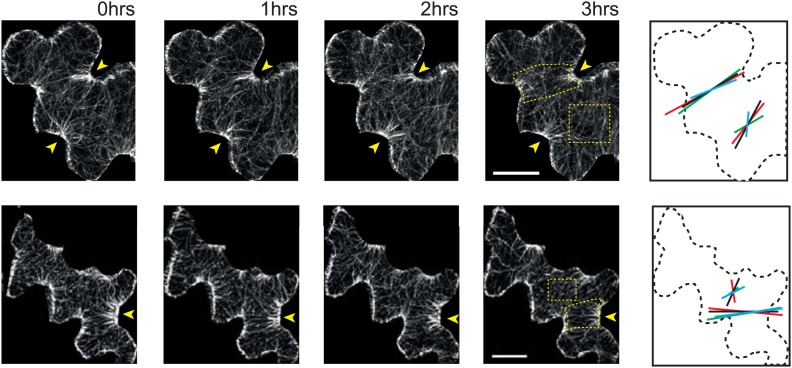
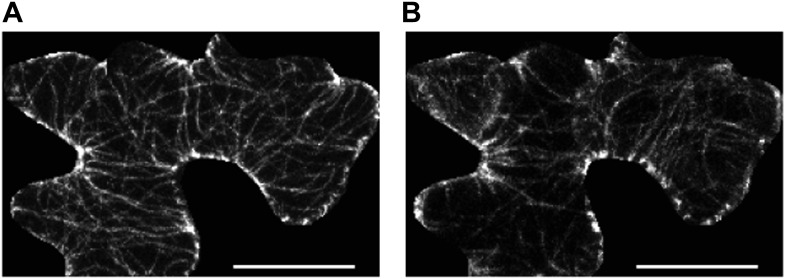
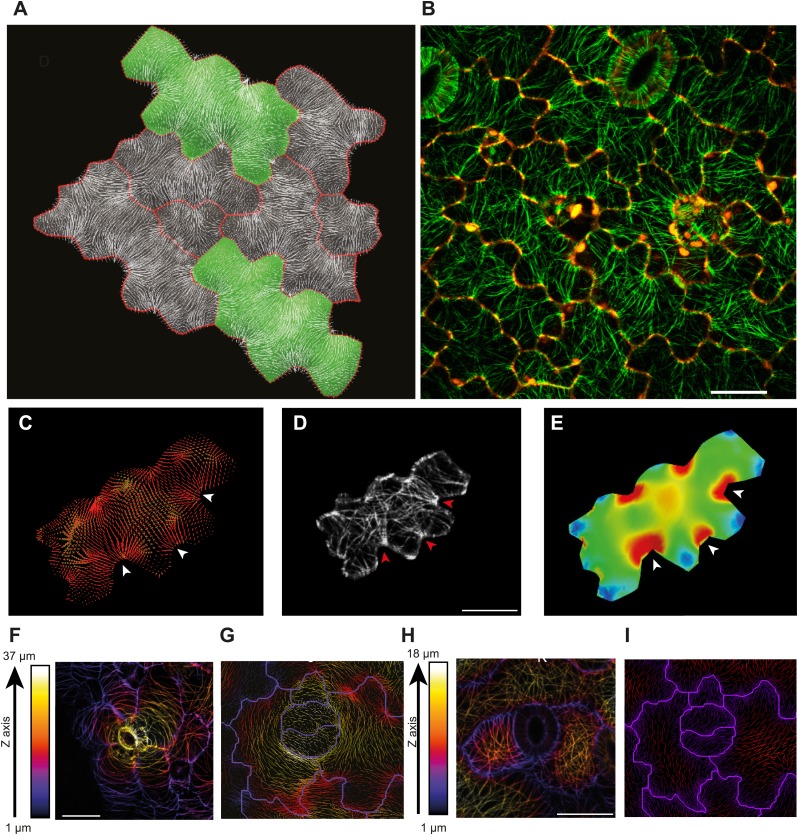
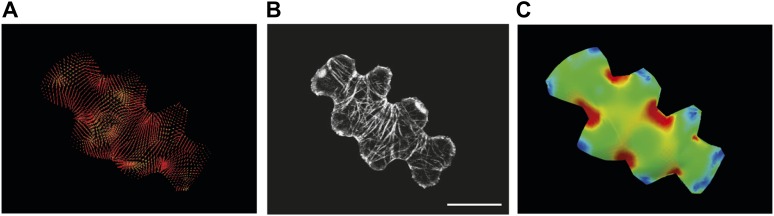
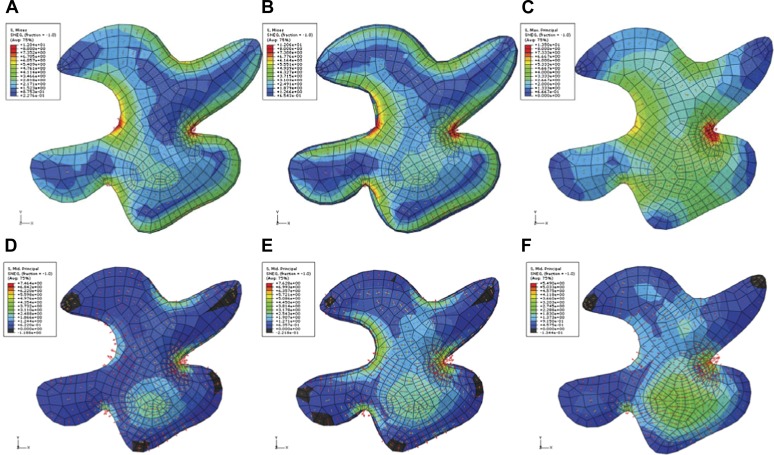
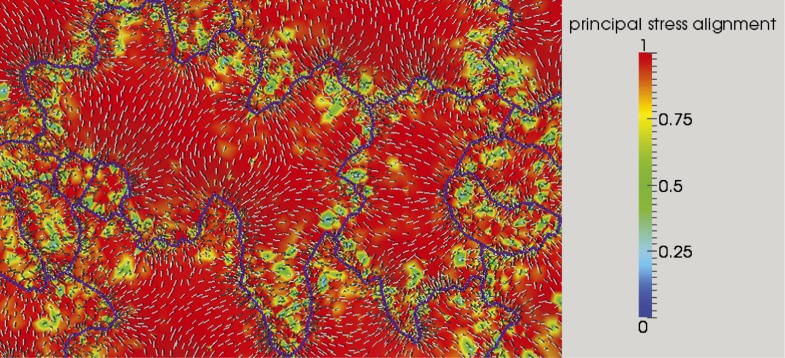
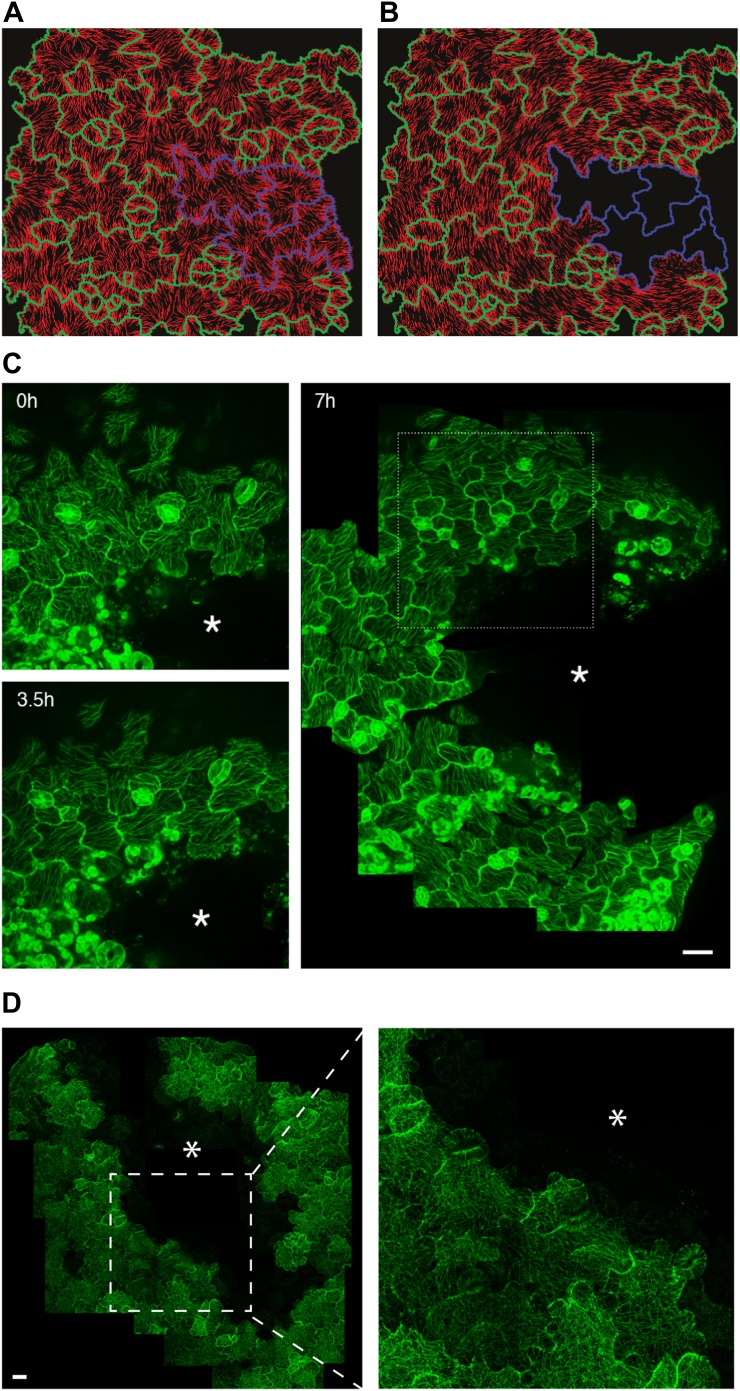
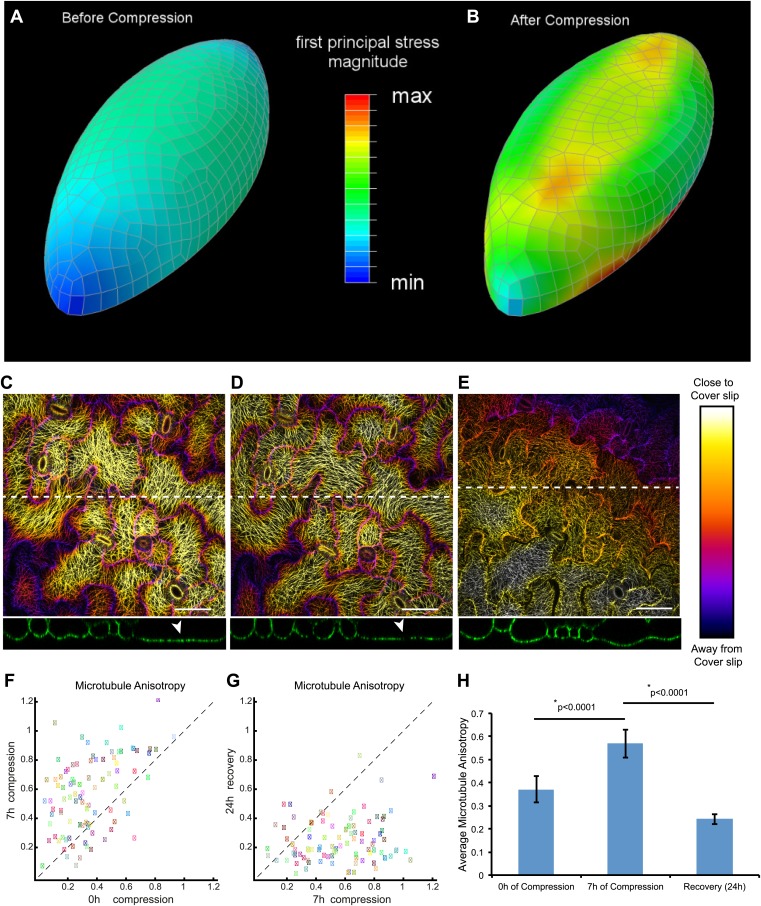
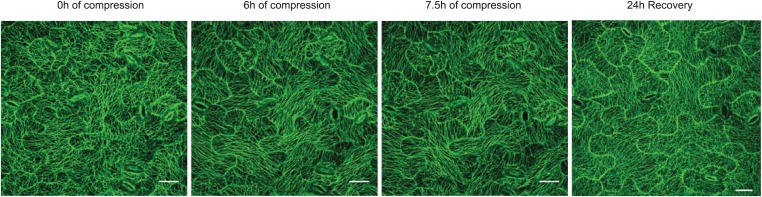
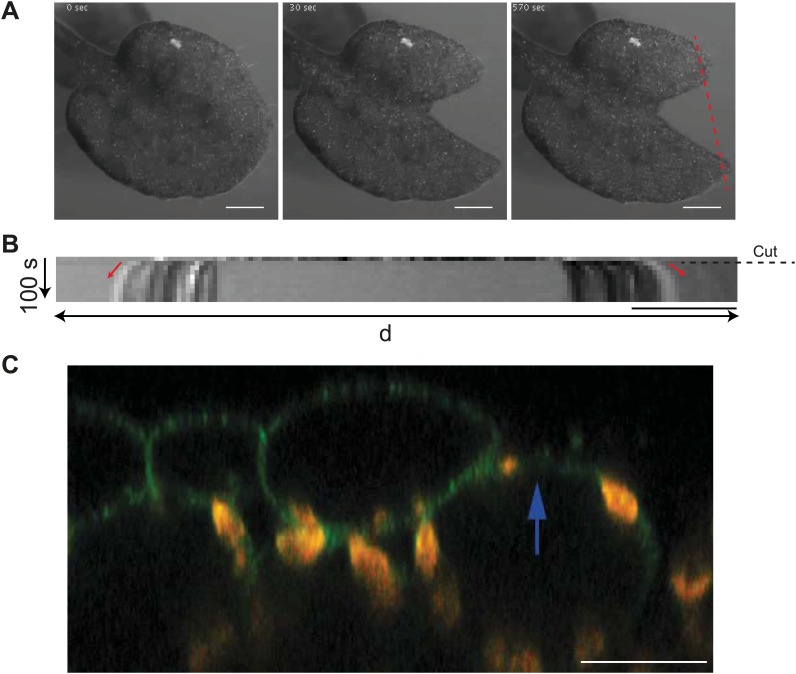
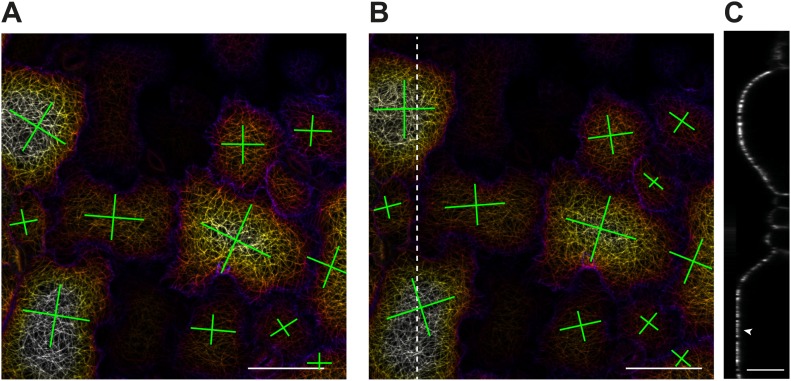
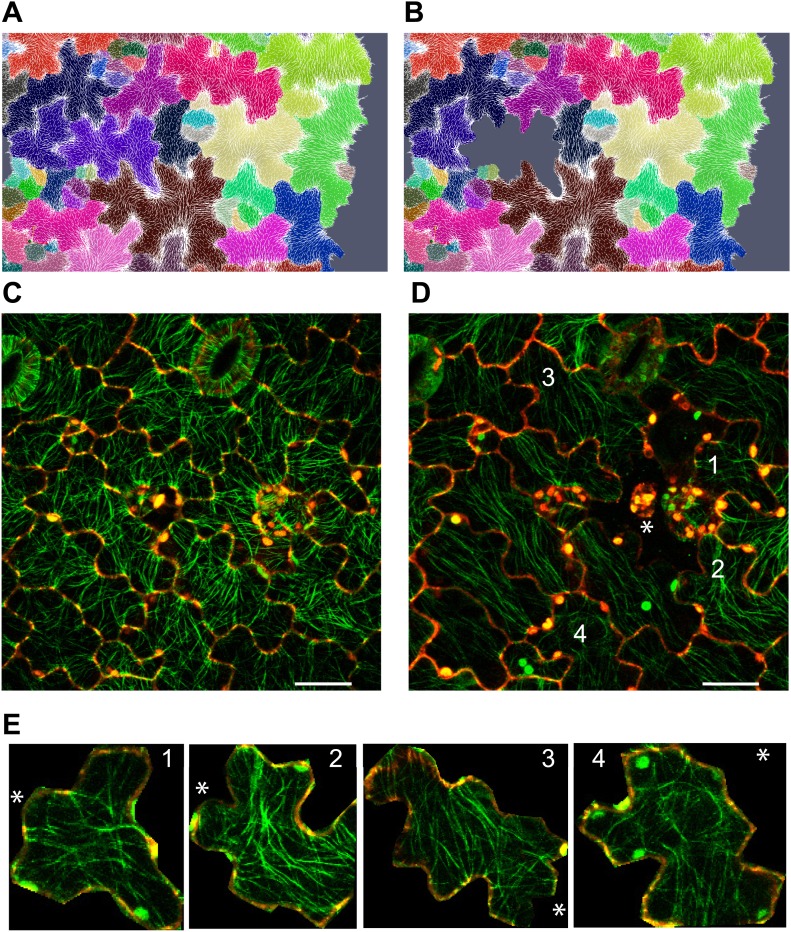
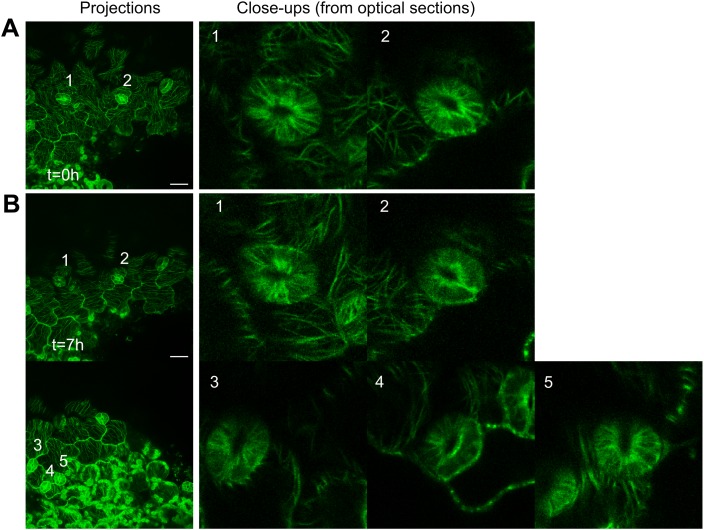
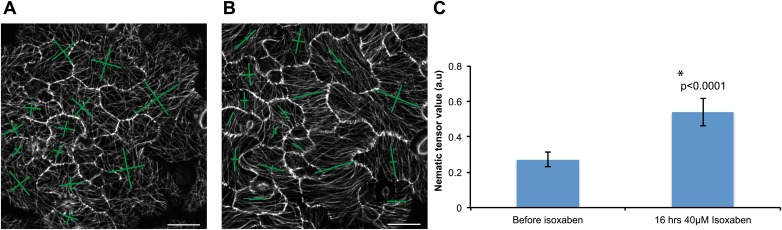
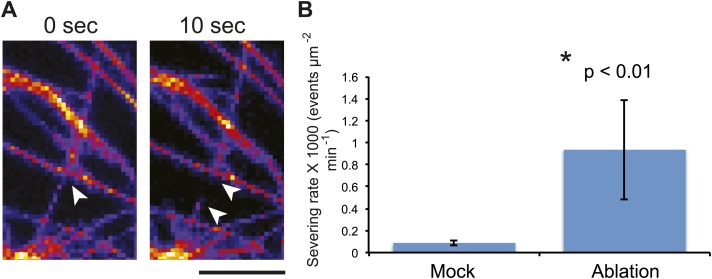
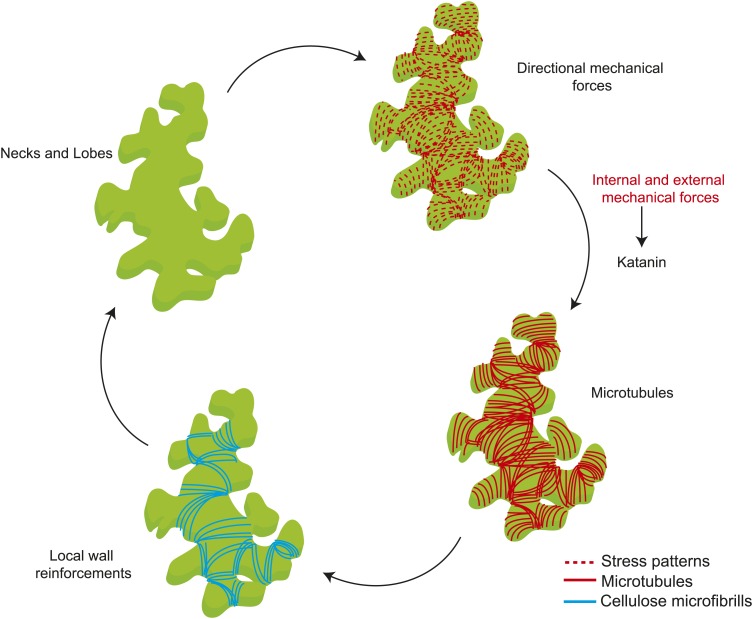
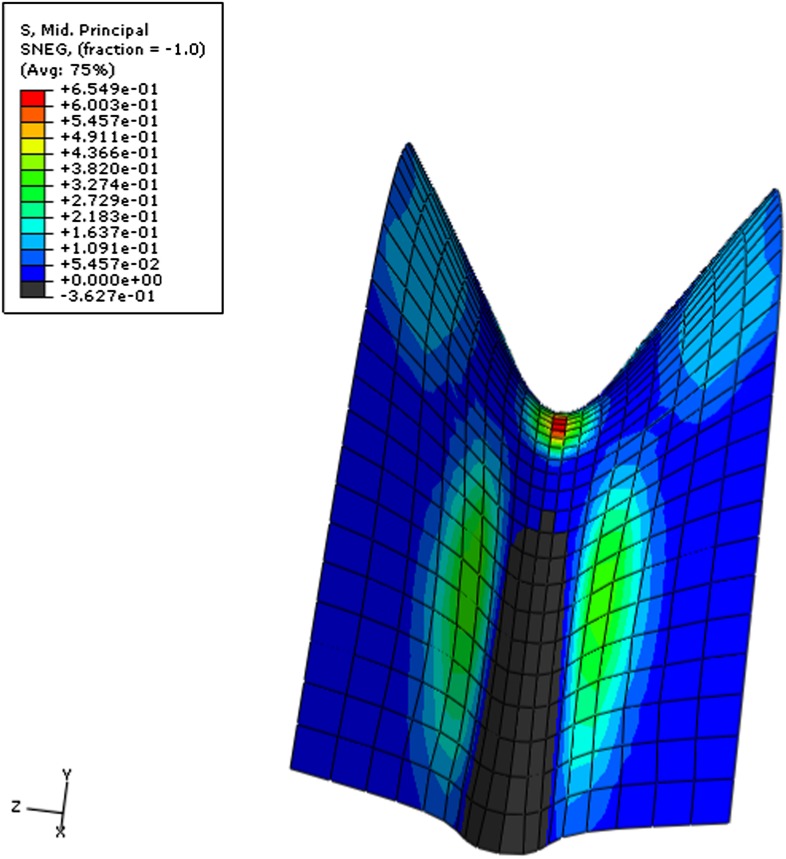
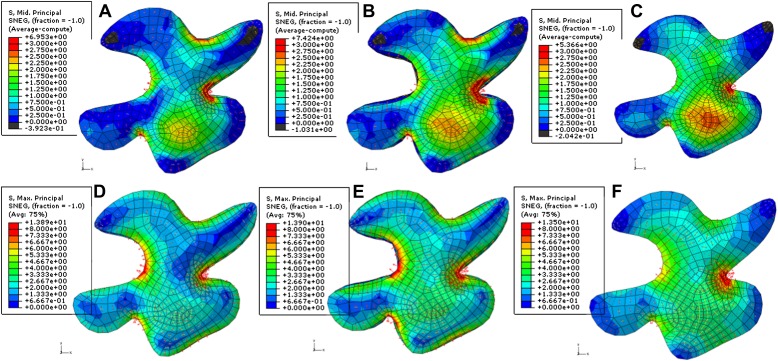
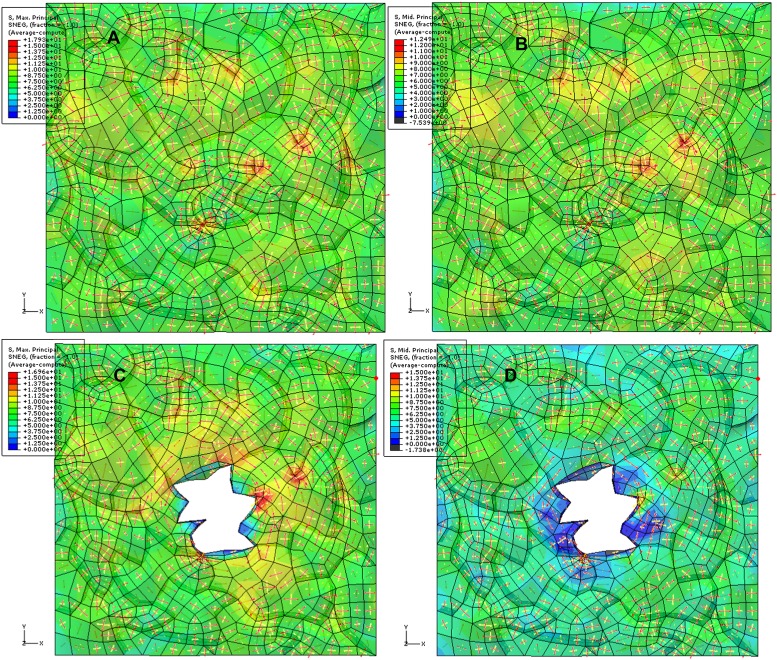
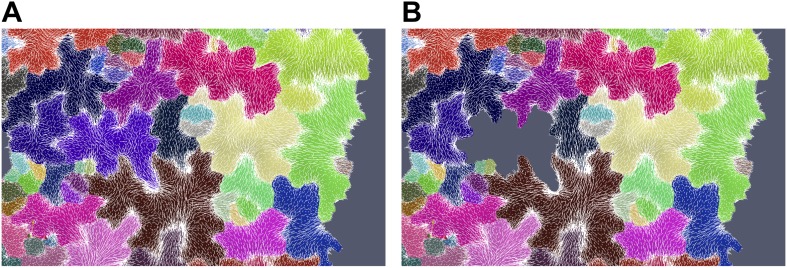
Although it is a central question in biology, how cell shape controls intracellular dynamics largely remains an open question. Here, we show that the shape of Arabidopsis pavement cells creates a stress pattern that controls microtubule orientation, which then guides cell wall reinforcement. Live-imaging, combined with modeling of cell mechanics, shows that microtubules align along the maximal tensile stress direction within the cells, and atomic force microscopy demonstrates that this leads to reinforcement of the cell wall parallel to the microtubules. This feedback loop is regulated: cell-shape derived stresses could be overridden by imposed tissue level stresses, showing how competition between subcellular and supracellular cues control microtubule behavior. Furthermore, at the microtubule level, we identified an amplification mechanism in which mechanical stress promotes the microtubule response to stress by increasing severing activity. These multiscale feedbacks likely contribute to the robustness of microtubule behavior in plant epidermis. DOI: http://dx.doi.org/10.7554/eLife.01967.001.
尽管这是生物学中的一个核心问题,但细胞形状如何控制细胞内动力学在很大程度上仍然是一个悬而未决的问题。在这里,我们表明拟南芥叶表皮细胞的形状会产生一种应力模式,该模式控制微管的方向,进而引导细胞壁的强化。实时成像与细胞力学建模相结合表明,微管沿着细胞内最大拉应力方向排列,原子力显微镜证明这会导致细胞壁平行于微管进行强化。这个反馈回路是受调控的:细胞形状衍生的应力可能会被施加的组织水平应力所覆盖,这表明亚细胞和超细胞信号之间的竞争如何控制微管行为。此外,在微管水平上,我们确定了一种放大机制,即机械应力通过增加切断活性来促进微管对应力的反应。这些多尺度反馈可能有助于植物表皮中微管行为的稳健性。DOI: http://dx.doi.org/10.7554/eLife.01967.001