Lee Richard W, Nicholson Lindsay B, Sen H Nida, Chan Chi-Chao, Wei Lai, Nussenblatt Robert B, Dick Andrew D
National Institute for Health Research Biomedical Research Centre at Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology, University Hospitals Bristol NHS, Foundation Trust, and University of Bristol, Bristol, UK.
Semin Immunopathol. 2014 Sep;36(5):581-94. doi: 10.1007/s00281-014-0433-9. Epub 2014 May 24.
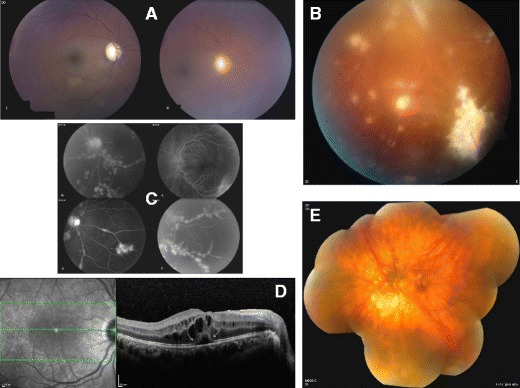
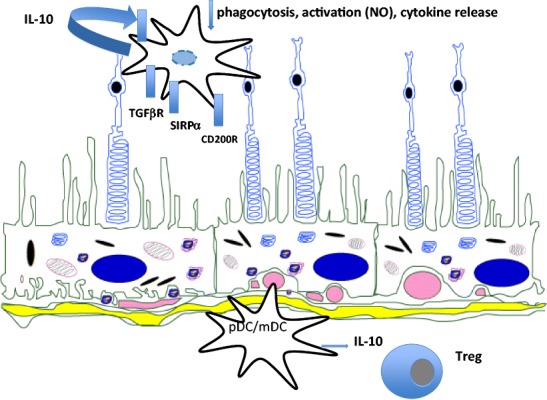
The eye, as currently viewed, is neither immunologically ignorant nor sequestered from the systemic environment. The eye utilises distinct immunoregulatory mechanisms to preserve tissue and cellular function in the face of immune-mediated insult; clinically, inflammation following such an insult is termed uveitis. The intra-ocular inflammation in uveitis may be clinically obvious as a result of infection (e.g. toxoplasma, herpes), but in the main infection, if any, remains covert. We now recognise that healthy tissues including the retina have regulatory mechanisms imparted by control of myeloid cells through receptors (e.g. CD200R) and soluble inhibitory factors (e.g. alpha-MSH), regulation of the blood retinal barrier, and active immune surveillance. Once homoeostasis has been disrupted and inflammation ensues, the mechanisms to regulate inflammation, including T cell apoptosis, generation of Treg cells, and myeloid cell suppression in situ, are less successful. Why inflammation becomes persistent remains unknown, but extrapolating from animal models, possibilities include differential trafficking of T cells from the retina, residency of CD8(+) T cells, and alterations of myeloid cell phenotype and function. Translating lessons learned from animal models to humans has been helped by system biology approaches and informatics, which suggest that diseased animals and people share similar changes in T cell phenotypes and monocyte function to date. Together the data infer a possible cryptic infectious drive in uveitis that unlocks and drives persistent autoimmune responses, or promotes further innate immune responses. Thus there may be many mechanisms in common with those observed in autoinflammatory disorders.
就目前的观点而言,眼睛既不是免疫忽视的,也并非与全身环境隔绝。面对免疫介导的损伤时,眼睛利用独特的免疫调节机制来维持组织和细胞功能;临床上,这种损伤后的炎症被称为葡萄膜炎。葡萄膜炎中的眼内炎症在感染(如弓形虫、疱疹)时临床上可能很明显,但在大多数情况下,感染(如果有)仍不明显。我们现在认识到,包括视网膜在内的健康组织具有通过受体(如CD200R)和可溶性抑制因子(如α-MSH)对髓样细胞进行控制、调节血视网膜屏障以及进行主动免疫监视所赋予的调节机制。一旦稳态被破坏并引发炎症,调节炎症的机制,包括T细胞凋亡、调节性T细胞的产生以及原位髓样细胞抑制,就不太成功了。炎症为何会持续仍然未知,但从动物模型推断,可能性包括T细胞从视网膜的差异迁移、CD8(+)T细胞的驻留以及髓样细胞表型和功能的改变。系统生物学方法和信息学有助于将从动物模型中学到的知识应用于人类,这表明到目前为止,患病动物和人类在T细胞表型和单核细胞功能方面有相似的变化。这些数据共同推断,葡萄膜炎中可能存在一种隐匿的感染驱动因素,它开启并驱动持续的自身免疫反应,或促进进一步的固有免疫反应。因此,可能存在许多与自身炎症性疾病中观察到的机制相同的机制。